

Mapping of foot-and-mouth disease virus antigenic sites recognized by single-domain antibodies reveals different 146S particle specific sites and particle flexibility
- PMID: 36699337
- PMCID: PMC9869066
- DOI: 10.3389/fvets.2022.1040802
Mapping of foot-and-mouth disease virus antigenic sites recognized by single-domain antibodies reveals different 146S particle specific sites and particle flexibility
Abstract
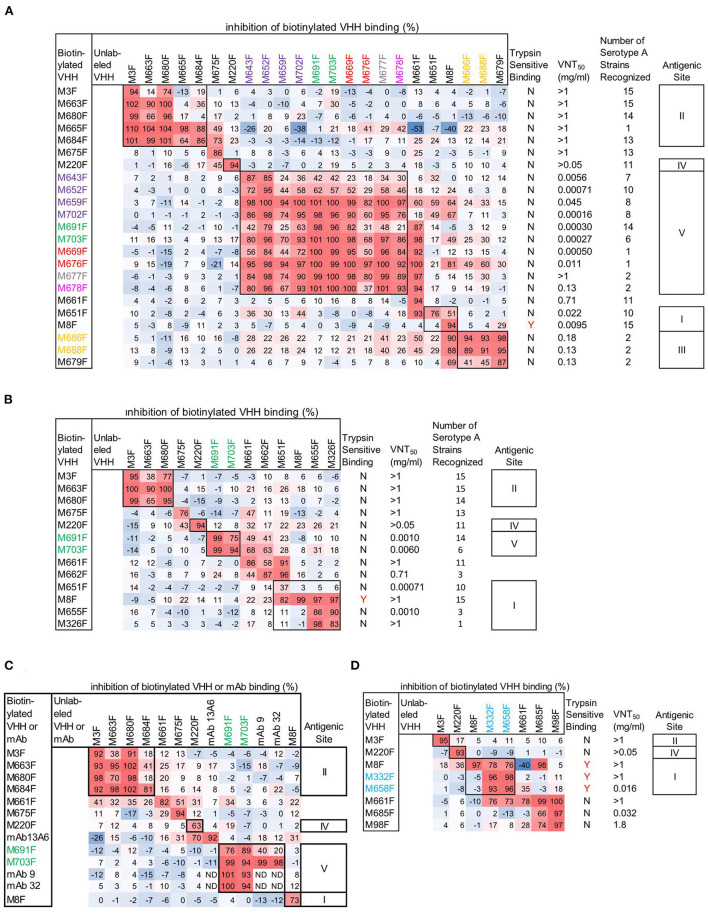
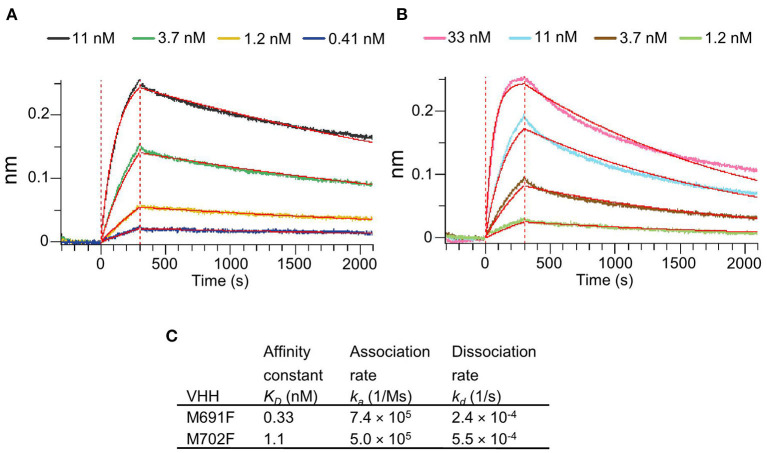
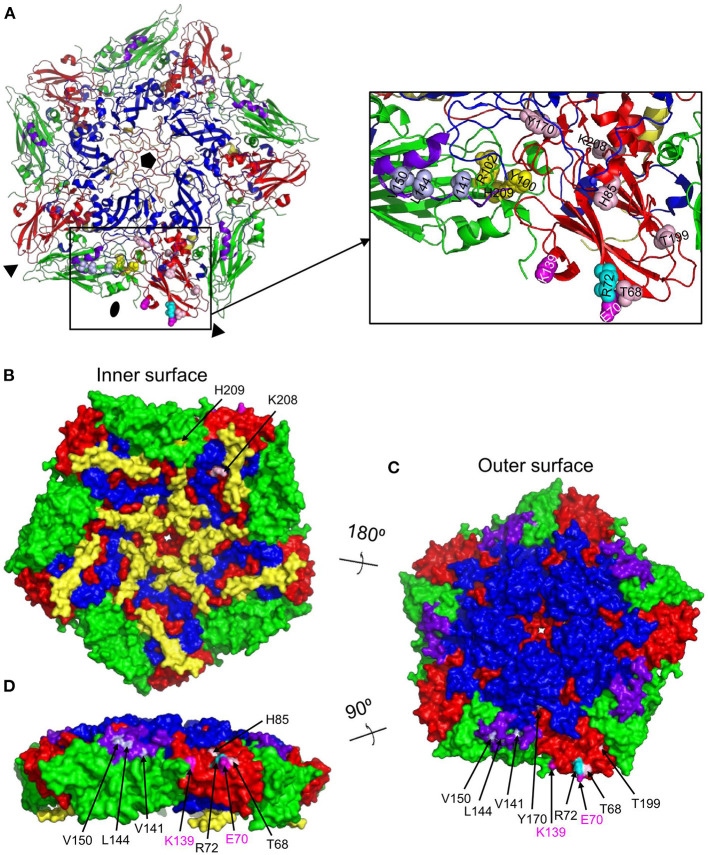
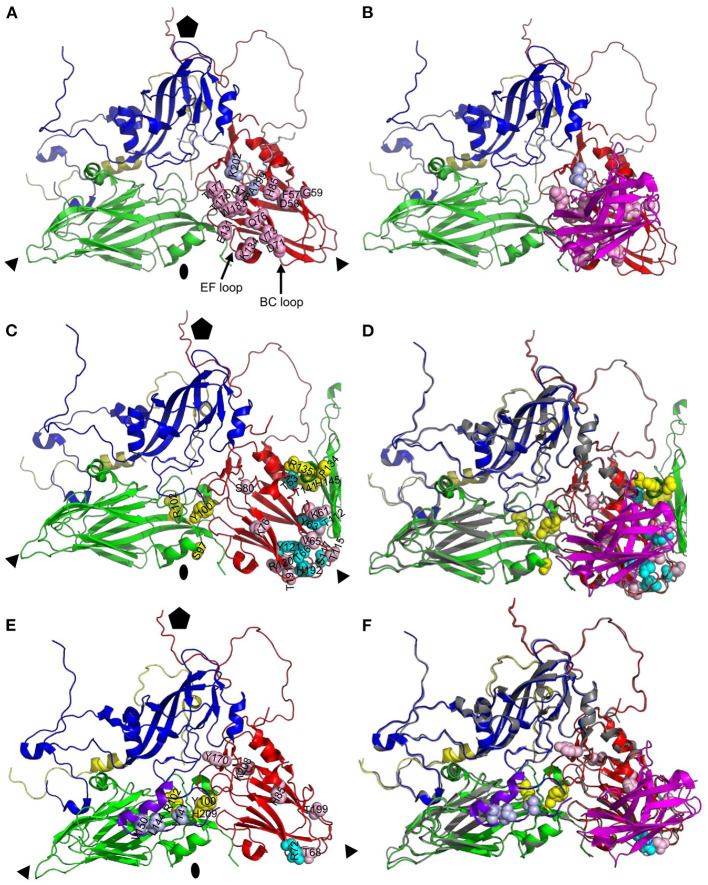
Vaccination with intact (146S) foot-and-mouth disease virus (FMDV) particles is used to control FMD. However, 146S particles easily dissociate into stable pentameric 12S particles which are less immunogenic. We earlier isolated several single-domain antibody fragments (VHHs) that specifically bind either 146S or 12S particles. These particle-specific VHHs are excellent tools for vaccine quality control. In this study we mapped the antigenic sites recognized by these VHHs by competition ELISAs, virus neutralization, and trypsin sensitivity of epitopes. We included two previously described monoclonal antibodies (mAbs) that are either 12S specific (mAb 13A6) or 146S specific (mAb 9). Although both are 12S specific, the VHH M3F and mAb 13A6 were found to bind independent antigenic sites. M3F recognized a non-neutralizing and trypsin insensitive site whereas mAb 13A6 recognized the trypsin sensitive VP2 N-terminus. The Asia1 146S-specific site was trypsin sensitive, neutralizing and also recognized by the VHH M8F, suggesting it involves the VP1 GH-loop. The type A 146S-specific VHHs recognized two independent antigenic sites that are both also neutralizing but trypsin insensitive. The major site was further mapped by cross-linking mass spectrometry (XL-MS) of two broadly strain reactive 146S-specific VHHs complexed to FMDV. The epitopes were located close to the 2-fold and 3-fold symmetry axes of the icosahedral virus 3D structure, mainly on VP2 and VP3, overlapping the earlier identified mAb 9 site. Since the epitopes were located on a single 12S pentamer, the 146S specificity cannot be explained by the epitope being split due to 12S pentamer dissociation. In an earlier study the cryo-EM structure of the 146S-specific VHH M170 complexed to type O FMDV was resolved. The 146S specificity was reported to be caused by an altered conformation of this epitope in 12S and 146S particles. This mechanism probably also explains the 146S-specific binding by the two type A VHHs mapped by XL-MS since their epitopes overlapped with the epitope recognized by M170. Surprisingly, residues internal in the 146S quaternary structure were also cross-linked to VHH. This probably reflects particle flexibility in solution. Molecular studies of virus-antibody interactions help to further optimize vaccines and improve their quality control.
Keywords: ELISA; VHH; XL-MS; epitope; foot-and-mouth disease virus (FMDV); nanobody; neutralizing antibody.
Copyright © 2023 Harmsen, Li, Sun, van der Poel and Dekker.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Single-Domain Antibodies That Specifically Recognize Intact Capsids of Multiple Foot-and-Mouth Disease Serotype O Strains.Vaccines (Basel). 2025 May 8;13(5):500. doi: 10.3390/vaccines13050500. Vaccines (Basel). 2025. PMID: 40432111 Free PMC article.
-
Isolation of Single-Domain Antibody Fragments That Preferentially Detect Intact (146S) Particles of Foot-and-Mouth Disease Virus for Use in Vaccine Quality Control.Front Immunol. 2017 Aug 17;8:960. doi: 10.3389/fimmu.2017.00960. eCollection 2017. Front Immunol. 2017. PMID: 28861080 Free PMC article.
-
Foot-and-mouth disease virus antigenic landscape and reduced immunogenicity elucidated in atomic detail.Nat Commun. 2024 Oct 10;15(1):8774. doi: 10.1038/s41467-024-53027-5. Nat Commun. 2024. PMID: 39389971 Free PMC article.
-
Single domain Camelid antibody fragments for molecular imaging and therapy of cancer.Front Oncol. 2023 Sep 8;13:1257175. doi: 10.3389/fonc.2023.1257175. eCollection 2023. Front Oncol. 2023. PMID: 37746282 Free PMC article. Review.
-
Promising Diagnostic and Therapeutic Approaches Based on VHHs for Cancer Management.Cancers (Basel). 2024 Jan 15;16(2):371. doi: 10.3390/cancers16020371. Cancers (Basel). 2024. PMID: 38254860 Free PMC article. Review.
Cited by
-
A broadly reactive ultralong bovine antibody that can determine the integrity of foot-and-mouth disease virus capsids.J Gen Virol. 2024 Oct;105(10):002032. doi: 10.1099/jgv.0.002032. J Gen Virol. 2024. PMID: 39422666 Free PMC article.
-
Serological Conversion through a Second Exposure to Inactivated Foot-and-Mouth Disease Virus Expressing the JC Epitope on the Viral Surface.Vaccines (Basel). 2023 Sep 14;11(9):1487. doi: 10.3390/vaccines11091487. Vaccines (Basel). 2023. PMID: 37766163 Free PMC article.
-
Cell Culture Adaptive Amino Acid Substitutions in FMDV Structural Proteins: A Key Mechanism for Altered Receptor Tropism.Viruses. 2024 Mar 27;16(4):512. doi: 10.3390/v16040512. Viruses. 2024. PMID: 38675855 Free PMC article. Review.
-
Single-Domain Antibodies That Specifically Recognize Intact Capsids of Multiple Foot-and-Mouth Disease Serotype O Strains.Vaccines (Basel). 2025 May 8;13(5):500. doi: 10.3390/vaccines13050500. Vaccines (Basel). 2025. PMID: 40432111 Free PMC article.
-
Conserved antigen structures and antibody-driven variations on foot-and-mouth disease virus serotype A revealed by bovine neutralizing monoclonal antibodies.PLoS Pathog. 2023 Nov 20;19(11):e1011811. doi: 10.1371/journal.ppat.1011811. eCollection 2023 Nov. PLoS Pathog. 2023. PMID: 37983290 Free PMC article.
References
-
- Mignaqui AC, Ferella A, Cass B, Mukankurayija L., L'Abbe D, Bisson L, et al. . Foot-and-mouth disease: optimization, reproducibility, and scalability of high-yield production of virus-like particles for a next-generation vaccine. Front Vet Sci. (2020) 7:601. 10.3389/fvets.2020.00601 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources