

Chronic Δ9-tetrahydrocannabinol impact on plasticity, and differential activation requirement for CB1-dependent long-term depression in ventral tegmental area GABA neurons in adult versus young mice
- PMID: 36699526
- PMCID: PMC9869137
- DOI: 10.3389/fnins.2022.1067493
Chronic Δ9-tetrahydrocannabinol impact on plasticity, and differential activation requirement for CB1-dependent long-term depression in ventral tegmental area GABA neurons in adult versus young mice
Abstract
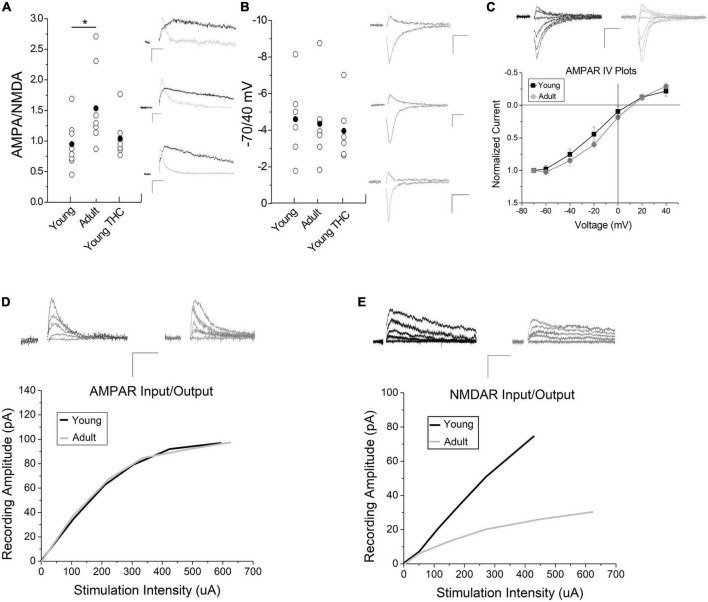
The ventral tegmental area (VTA) mediates incentive salience and reward prediction error through dopamine (DA) neurons that are regulated by local VTA GABA neurons. In young mice, VTA GABA cells exhibit a form of synaptic plasticity known as long-term depression (LTD) that is dependent on cannabinoid 1 (CB1) receptors preceded by metabotropic glutamate receptor 5 (mGluR5) signaling to induce endocannabinoid production. This LTD was eliminated following chronic (7-10 consecutive days) exposure to the marijuana derived cannabinoid Δ9 -tetrahydrocannabinol (THC). We now examine the mechanism behind THC-induced elimination of LTD in adolescents as well as plasticity induction ability in adult versus young male and female mice using whole-cell electrophysiology experiments of VTA GABA cells. Chronic THC injections in adolescents resulted in a loss of CB1 agonist-mediated depression, illustrating chronic THC likely desensitizes or removes synaptic CB1. We noted that seven days withdrawal from chronic THC restored LTD and CB1 agonist-induced depression, suggesting reversibility of THC-induced changes. Adult mice continue to express functional mGluR5 and CB1, but require a doubling of the synaptic stimulation compared to young mice to induce LTD, suggesting a quantitative difference in CB1-dependent plasticity between young and adult mice. One potential rationale for this difference is changes in AMPA and NMDA glutamate receptors. Indeed, AMPA/NMDA ratios were increased in in adults compared to young mice. Lastly, we performed quantitative reverse-transcription PCR and identified that CB1, DAGLα, and GluA1 levels increased following chronic THC exposure. Collectively, our data demonstrate the first age-dependent GABA neuron plasticity in the VTA, which could have implications for decreased THC dependence capacity in adults, as well as the mechanism behind chronic THC-induced synaptic alterations in young mice.
Keywords: CB1; GABA; LTD; THC; VTA; age; development.
Copyright © 2023 Ostlund, Von Gunten, Smith and Edwards.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
Three-day delta-9-tetrahydrocannabinol (THC) exposure eliminates long-term depression in ventral tegmental area of young, but not adult mice.J Cannabis Res. 2025 May 31;7(1):30. doi: 10.1186/s42238-025-00287-7. J Cannabis Res. 2025. PMID: 40450354 Free PMC article.
-
CB1-Dependent Long-Term Depression in Ventral Tegmental Area GABA Neurons: A Novel Target for Marijuana.J Neurosci. 2017 Nov 8;37(45):10943-10954. doi: 10.1523/JNEUROSCI.0190-17.2017. Epub 2017 Oct 16. J Neurosci. 2017. PMID: 29038246 Free PMC article.
-
Acute cocaine exposure occludes long-term depression in ventral tegmental area GABA neurons.Neurochem Int. 2021 May;145:105002. doi: 10.1016/j.neuint.2021.105002. Epub 2021 Feb 20. Neurochem Int. 2021. PMID: 33617930 Free PMC article.
-
New perspectives in the studies on endocannabinoid and cannabis: abnormal behaviors associate with CB1 cannabinoid receptor and development of therapeutic application.J Pharmacol Sci. 2004 Dec;96(4):362-6. doi: 10.1254/jphs.fmj04003x2. Epub 2004 Dec 10. J Pharmacol Sci. 2004. PMID: 15599103 Review.
-
[Epigenetic effects of cannabis/tetrahydrocannabinol].Bull Acad Natl Med. 2020 Jun;204(6):570-576. doi: 10.1016/j.banm.2020.04.004. Epub 2020 Apr 15. Bull Acad Natl Med. 2020. PMID: 32296244 Free PMC article. Review. French.
Cited by
-
In silico analyses of the involvement of GPR55, CB1R and TRPV1: response to THC, contribution to temporal lobe epilepsy, structural modeling and updated evolution.Front Neuroinform. 2024 Feb 7;18:1294939. doi: 10.3389/fninf.2024.1294939. eCollection 2024. Front Neuroinform. 2024. PMID: 38404644 Free PMC article.
-
Cannabis, Endocannabinoids and Brain Development: From Embryogenesis to Adolescence.Cells. 2024 Nov 13;13(22):1875. doi: 10.3390/cells13221875. Cells. 2024. PMID: 39594623 Free PMC article. Review.
-
Automated neonatal nnU-Net brain MRI extractor trained on a large multi-institutional dataset.Sci Rep. 2024 Feb 26;14(1):4583. doi: 10.1038/s41598-024-54436-8. Sci Rep. 2024. PMID: 38403673 Free PMC article.
-
Genetic effect on working memory activation pattern and structural properties of cortical gray matter.Sci Rep. 2025 Jul 1;15(1):21096. doi: 10.1038/s41598-025-07959-7. Sci Rep. 2025. PMID: 40596557 Free PMC article.
-
Three-day delta-9-tetrahydrocannabinol (THC) exposure eliminates long-term depression in ventral tegmental area of young, but not adult mice.J Cannabis Res. 2025 May 31;7(1):30. doi: 10.1186/s42238-025-00287-7. J Cannabis Res. 2025. PMID: 40450354 Free PMC article.
References
-
- Al-Hasani R., Gowrishankar R., Schmitz G. P., Pedersen C. E., Marcus D. J., Shirley S. E., et al. (2021). Ventral tegmental area Gabaergic inhibition of cholinergic interneurons in the ventral nucleus accumbens shell promotes reward reinforcement. Nat. Neurosci. 24:1501. 10.1038/s41593-021-00928-z - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources