

Combined metabolomic and transcriptomic analysis reveals key components of OsCIPK17 overexpression improves drought tolerance in rice
- PMID: 36699859
- PMCID: PMC9868928
- DOI: 10.3389/fpls.2022.1043757
Combined metabolomic and transcriptomic analysis reveals key components of OsCIPK17 overexpression improves drought tolerance in rice
Abstract
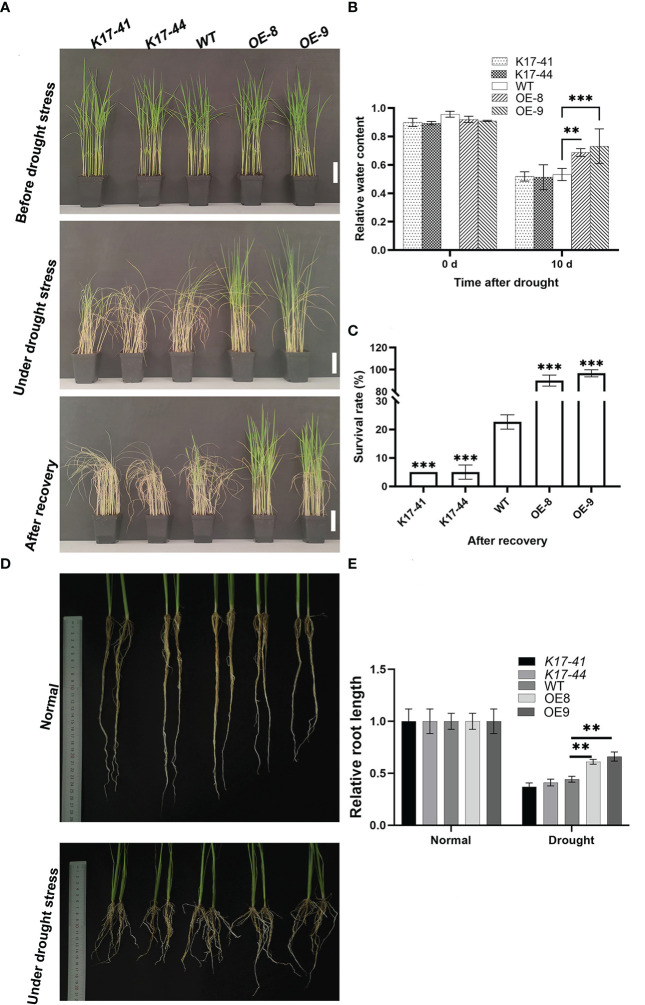
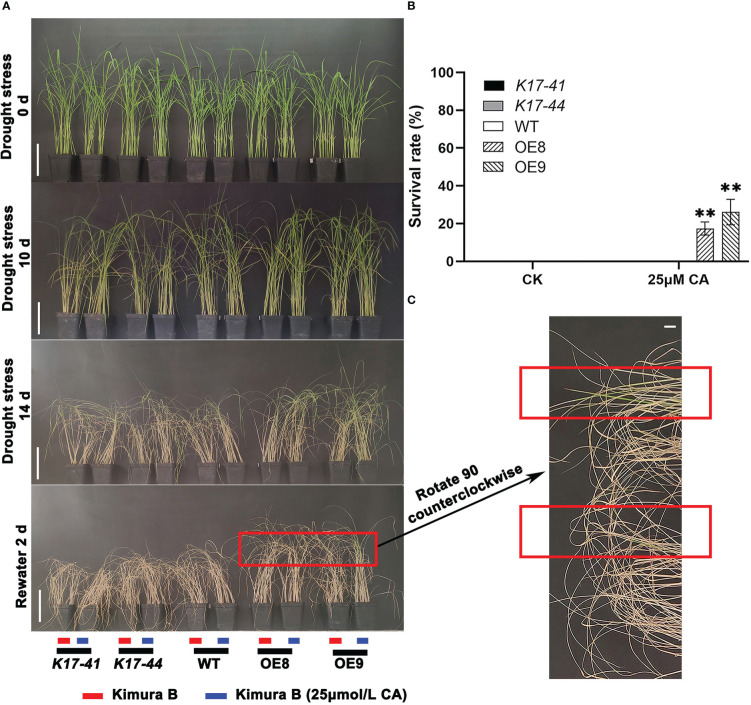
Oryza Sativa is one of the most important food crops in China, which is easily affected by drought during its growth and development. As a member of the calcium signaling pathway, CBL-interacting protein kinase (CIPK) plays an important role in plant growth and development as well as environmental stress. However, there is no report on the function and mechanism of OsCIPK17 in rice drought resistance. We combined transcriptional and metabonomic analysis to clarify the specific mechanism of OsCIPK17 in response to rice drought tolerance. The results showed that OsCIPK17 improved drought resistance of rice by regulating deep roots under drought stress; Response to drought by regulating the energy metabolism pathway and controlling the accumulation of citric acid in the tricarboxylic acid (TCA) cycle; Our exogenous experiments also proved that OsCIPK17 responds to citric acid, and this process involves the auxin metabolism pathway; Exogenous citric acid can improve the drought resistance of overexpression plants. Our research reveals that OsCIPK17 positively regulates rice drought resistance and participates in the accumulation of citric acid in the TCA cycle, providing new insights for rice drought resistance.
Keywords: CIPK; Oryza sativa; auxin; citric acid; drought; metabolome; transcriptome.
Copyright © 2023 Lu, Chen, Wang, Han, Zhao, Xue, Zhang, Fang, Wang and Cao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures








Similar articles
-
The OsCBL8-OsCIPK17 Module Regulates Seedling Growth and Confers Resistance to Heat and Drought in Rice.Int J Mol Sci. 2022 Oct 18;23(20):12451. doi: 10.3390/ijms232012451. Int J Mol Sci. 2022. PMID: 36293306 Free PMC article.
-
Combined Metabolomic and Transcriptomic Analysis Reveals Allantoin Enhances Drought Tolerance in Rice.Int J Mol Sci. 2022 Nov 16;23(22):14172. doi: 10.3390/ijms232214172. Int J Mol Sci. 2022. PMID: 36430648 Free PMC article.
-
Roles of a maize phytochrome-interacting factors protein ZmPIF3 in regulation of drought stress responses by controlling stomatal closure in transgenic rice without yield penalty.Plant Mol Biol. 2018 Jul;97(4-5):311-323. doi: 10.1007/s11103-018-0739-4. Epub 2018 Jun 5. Plant Mol Biol. 2018. PMID: 29869742
-
The soybean CBL-interacting protein kinase, GmCIPK2, positively regulates drought tolerance and ABA signaling.Plant Physiol Biochem. 2021 Oct;167:980-989. doi: 10.1016/j.plaphy.2021.09.026. Epub 2021 Sep 24. Plant Physiol Biochem. 2021. PMID: 34583133
-
Progress studies of drought-responsive genes in rice.Plant Cell Rep. 2011 Mar;30(3):297-310. doi: 10.1007/s00299-010-0956-z. Epub 2010 Dec 4. Plant Cell Rep. 2011. PMID: 21132431 Review.
Cited by
-
Physiological and Molecular Mechanisms of Rice Tolerance to Salt and Drought Stress: Advances and Future Directions.Int J Mol Sci. 2024 Aug 29;25(17):9404. doi: 10.3390/ijms25179404. Int J Mol Sci. 2024. PMID: 39273349 Free PMC article. Review.
-
Advances in Understanding Drought Stress Responses in Rice: Molecular Mechanisms of ABA Signaling and Breeding Prospects.Genes (Basel). 2024 Nov 27;15(12):1529. doi: 10.3390/genes15121529. Genes (Basel). 2024. PMID: 39766796 Free PMC article. Review.
-
OsCIPK2 mediated rice root microorganisms and metabolites to improve plant nitrogen uptake.BMC Plant Biol. 2024 Apr 16;24(1):285. doi: 10.1186/s12870-024-04982-0. BMC Plant Biol. 2024. PMID: 38627617 Free PMC article.
-
Integrated analysis of transcriptomics and metabolomics of garden asparagus (Asparagus officinalis L.) under drought stress.BMC Plant Biol. 2024 Jun 15;24(1):563. doi: 10.1186/s12870-024-05286-z. BMC Plant Biol. 2024. PMID: 38879466 Free PMC article.
-
Diethyl aminoethyl hexanoate ameliorates salt tolerance associated with ion transport, osmotic adjustment, and metabolite reprograming in white clover.BMC Plant Biol. 2024 Oct 12;24(1):950. doi: 10.1186/s12870-024-05657-6. BMC Plant Biol. 2024. PMID: 39394568 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous