

TadA orthologs enable both cytosine and adenine editing of base editors
- PMID: 36702837
- PMCID: PMC9880001
- DOI: 10.1038/s41467-023-36003-3
TadA orthologs enable both cytosine and adenine editing of base editors
Abstract
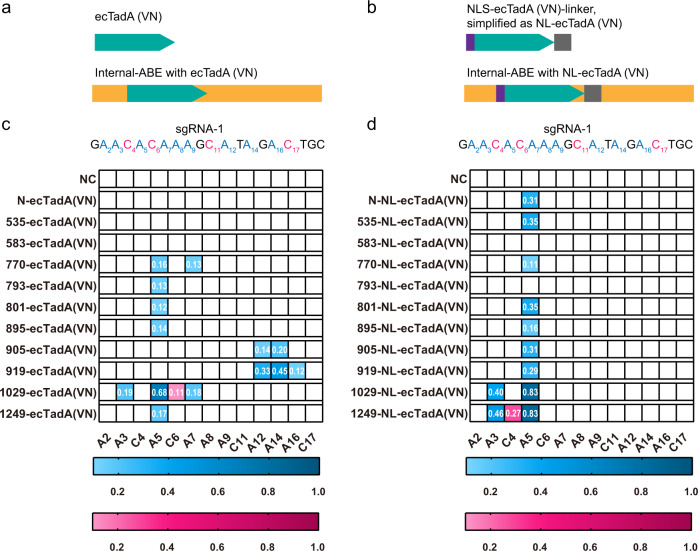
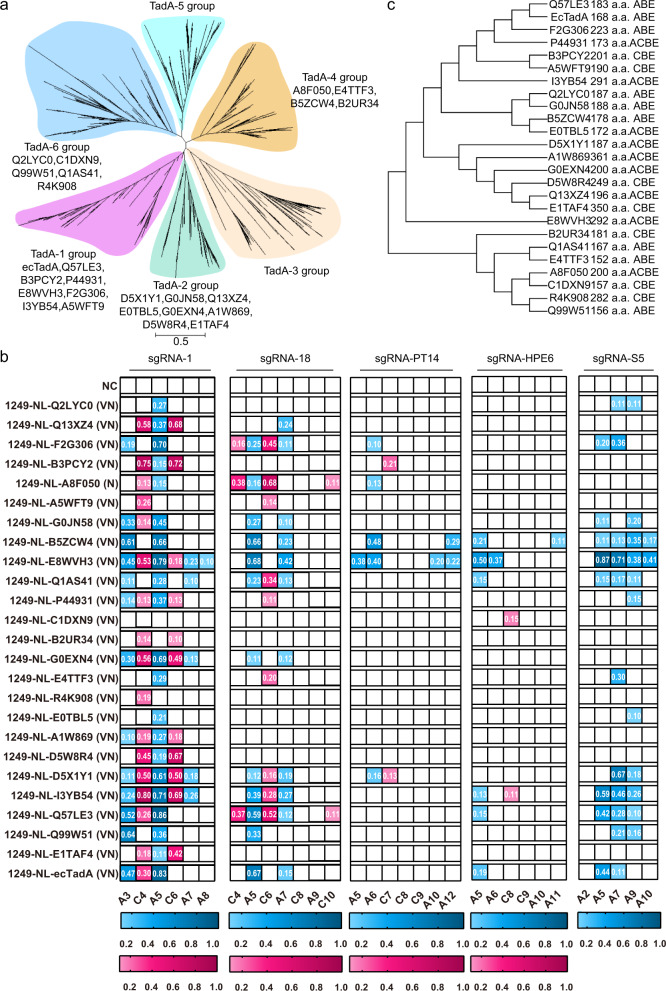
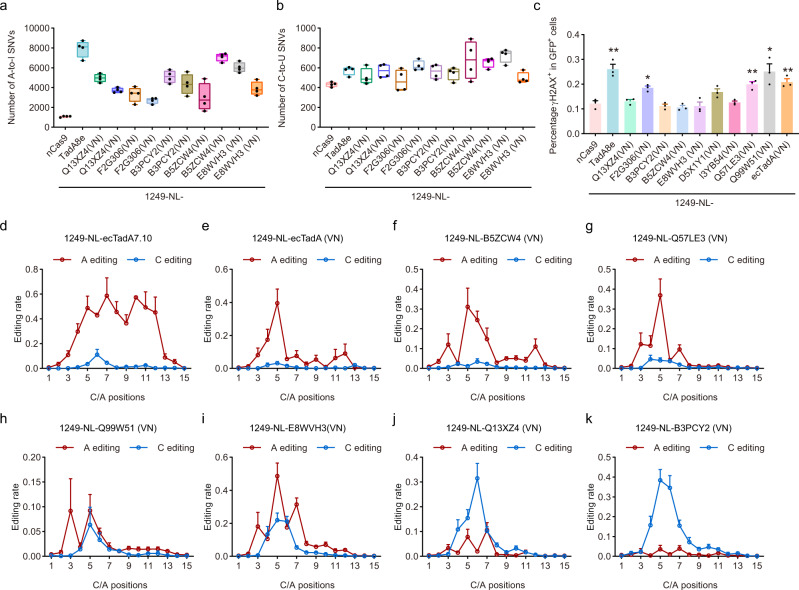
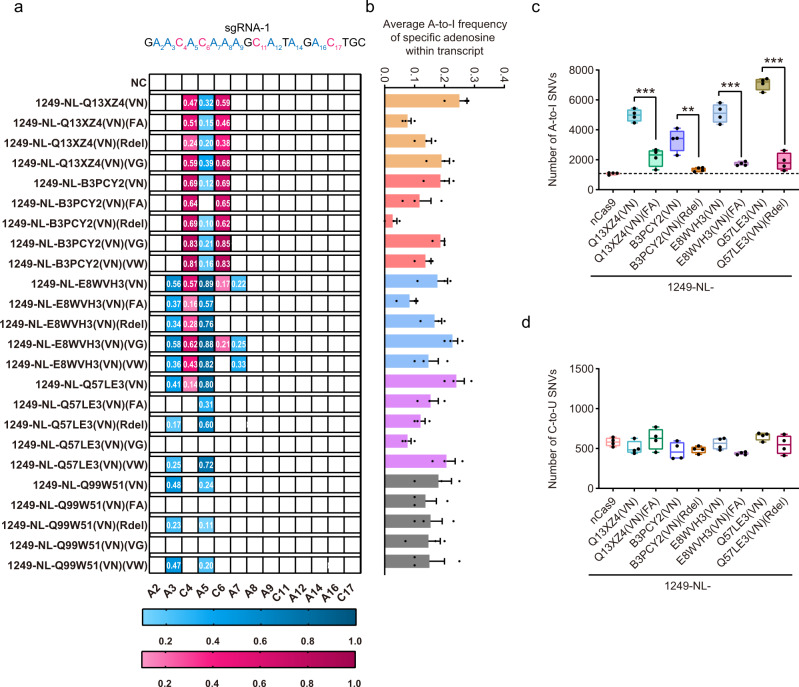
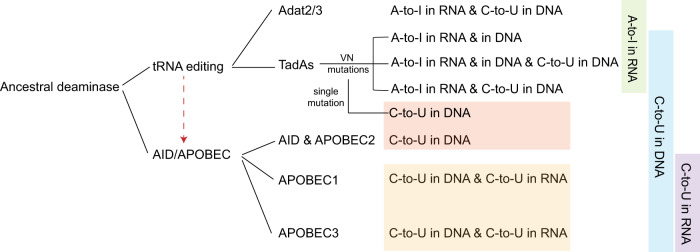
Cytidine and adenosine deaminases are required for cytosine and adenine editing of base editors respectively, and no single deaminase could enable concurrent and comparable cytosine and adenine editing. Additionally, distinct properties of cytidine and adenosine deaminases lead to various types of off-target effects, including Cas9-indendepent DNA off-target effects for cytosine base editors (CBEs) and RNA off-target effects particularly severe for adenine base editors (ABEs). Here we demonstrate that 25 TadA orthologs could be engineered to generate functional ABEs, CBEs or ACBEs via single or double mutations, which display minimized Cas9-independent DNA off-target effects and genotoxicity, with orthologs B5ZCW4, Q57LE3, E8WVH3, Q13XZ4 and B3PCY2 as promising candidates for further engineering. Furthermore, RNA off-target effects of TadA ortholog-derived base editors could be further reduced or even eliminated by additional single mutation. Taken together, our work expands the base editing toolkits, and also provides important clues for the potential evolutionary process of deaminases.
© 2023. The Author(s).
Conflict of interest statement
Fudan University has a patent (Chinese Patent Application No. 202111226226.X) pending, with T.-L.C., S.Z., and J.Y.Q. as inventors, for TadA orthologs described in this paper. The remaining authors declare no competing interests.
Figures
Similar articles
-
Evolution of an adenine base editor into a small, efficient cytosine base editor with low off-target activity.Nat Biotechnol. 2023 May;41(5):673-685. doi: 10.1038/s41587-022-01533-6. Epub 2022 Nov 10. Nat Biotechnol. 2023. PMID: 36357719 Free PMC article.
-
Improved cytosine base editors generated from TadA variants.Nat Biotechnol. 2023 May;41(5):686-697. doi: 10.1038/s41587-022-01611-9. Epub 2023 Jan 9. Nat Biotechnol. 2023. PMID: 36624149 Free PMC article.
-
Re-engineering the adenine deaminase TadA-8e for efficient and specific CRISPR-based cytosine base editing.Nat Biotechnol. 2023 May;41(5):663-672. doi: 10.1038/s41587-022-01532-7. Epub 2022 Nov 10. Nat Biotechnol. 2023. PMID: 36357717
-
Precision genome engineering through adenine and cytosine base editing.Nat Plants. 2018 Mar;4(3):148-151. doi: 10.1038/s41477-018-0115-z. Epub 2018 Feb 26. Nat Plants. 2018. PMID: 29483683 Review.
-
Off-Target Editing by CRISPR-Guided DNA Base Editors.Biochemistry. 2019 Sep 10;58(36):3727-3734. doi: 10.1021/acs.biochem.9b00573. Epub 2019 Aug 26. Biochemistry. 2019. PMID: 31433621 Free PMC article. Review.
Cited by
-
Advancing CRISPR base editing technology through innovative strategies and ideas.Sci China Life Sci. 2025 Mar;68(3):610-627. doi: 10.1007/s11427-024-2699-5. Epub 2024 Sep 2. Sci China Life Sci. 2025. PMID: 39231901 Review.
-
Charting the development and engineering of CRISPR base editors: lessons and inspirations.Cell Chem Biol. 2025 Jun 19;32(6):789-808. doi: 10.1016/j.chembiol.2025.05.003. Epub 2025 Jun 5. Cell Chem Biol. 2025. PMID: 40480225 Review.
-
Engineering TadA ortholog-derived cytosine base editor without motif preference and adenosine activity limitation.Nat Commun. 2024 Sep 16;15(1):8090. doi: 10.1038/s41467-024-52485-1. Nat Commun. 2024. PMID: 39284833 Free PMC article.
-
High-precision cytosine base editors by evolving nucleic-acid-recognition hotspots in deaminase.Nat Biotechnol. 2025 Jul 7:10.1038/s41587-025-02678-w. doi: 10.1038/s41587-025-02678-w. Online ahead of print. Nat Biotechnol. 2025. PMID: 40624252
-
Efforts to Downsize Base Editors for Clinical Applications.Int J Mol Sci. 2025 Mar 6;26(5):2357. doi: 10.3390/ijms26052357. Int J Mol Sci. 2025. PMID: 40076976 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
