

Rapid upwards spread of non-native plants in mountains across continents
- PMID: 36702858
- PMCID: PMC9998268
- DOI: 10.1038/s41559-022-01979-6
Rapid upwards spread of non-native plants in mountains across continents
Abstract
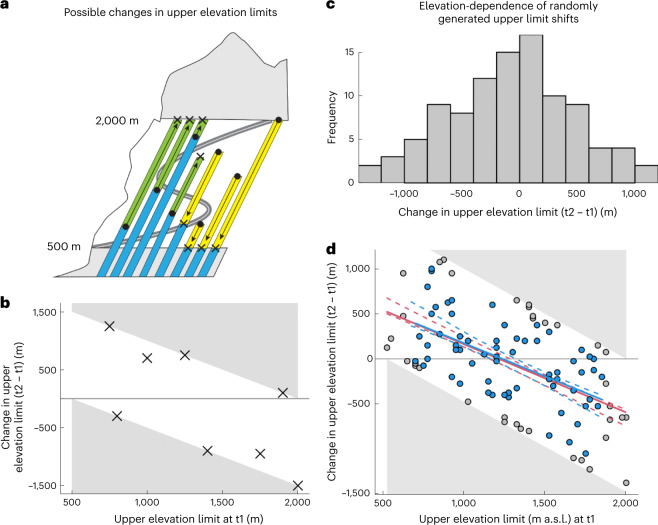
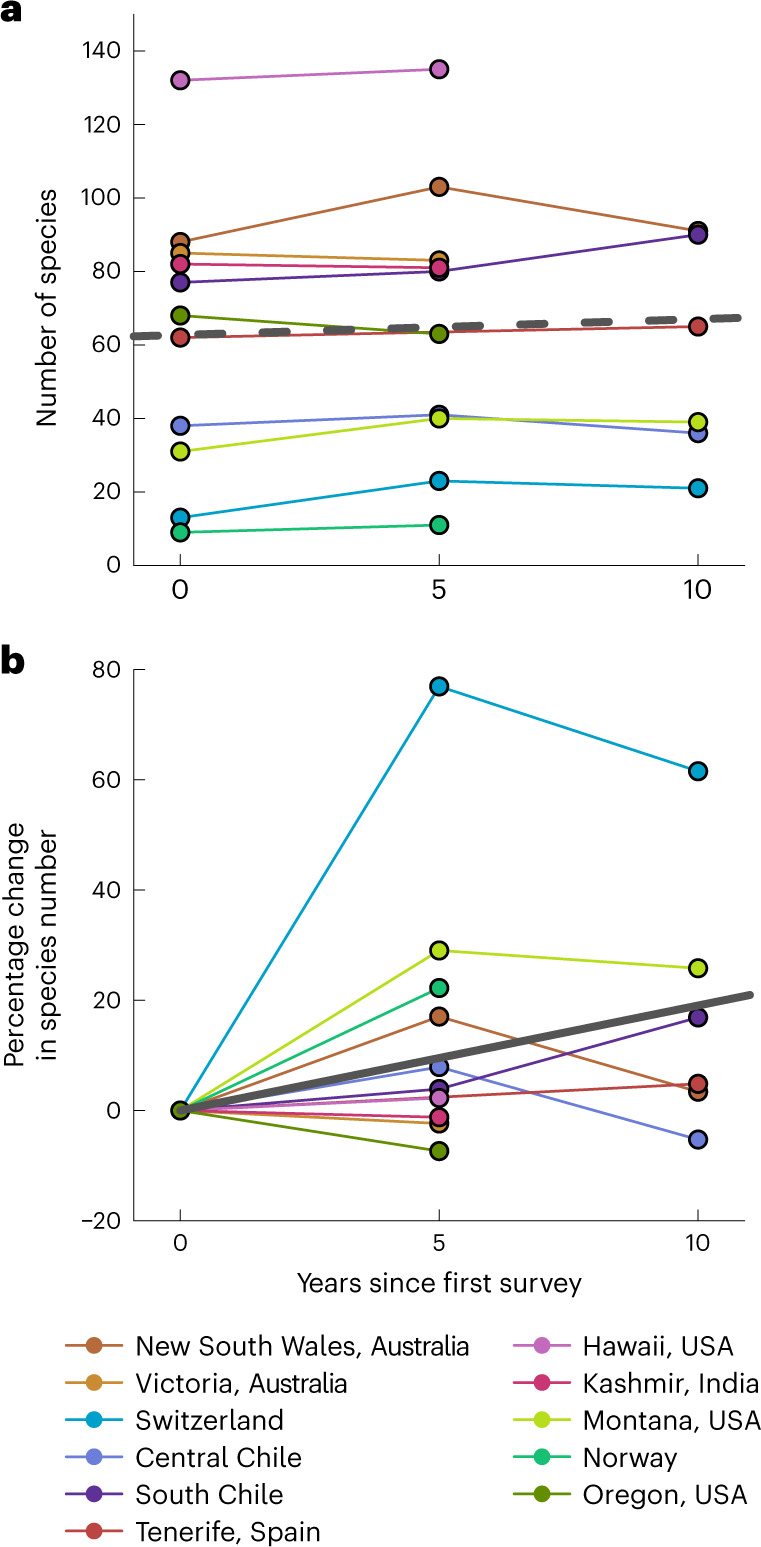
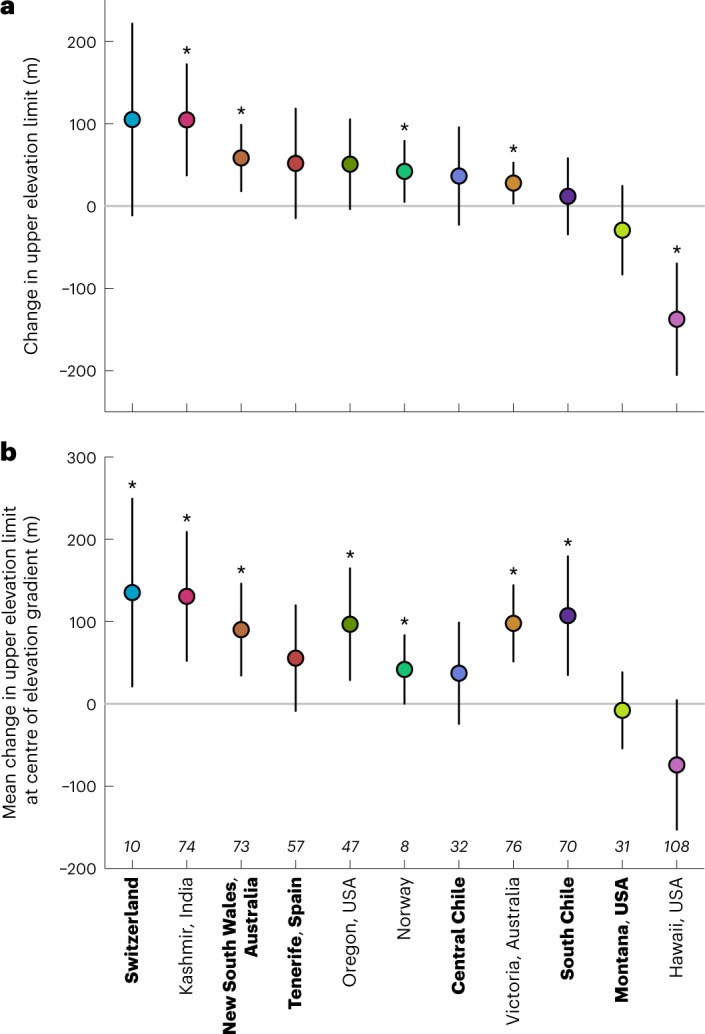
High-elevation ecosystems are among the few ecosystems worldwide that are not yet heavily invaded by non-native plants. This is expected to change as species expand their range limits upwards to fill their climatic niches and respond to ongoing anthropogenic disturbances. Yet, whether and how quickly these changes are happening has only been assessed in a few isolated cases. Starting in 2007, we conducted repeated surveys of non-native plant distributions along mountain roads in 11 regions from 5 continents. We show that over a 5- to 10-year period, the number of non-native species increased on average by approximately 16% per decade across regions. The direction and magnitude of upper range limit shifts depended on elevation across all regions. Supported by a null-model approach accounting for range changes expected by chance alone, we found greater than expected upward shifts at lower/mid elevations in at least seven regions. After accounting for elevation dependence, significant average upward shifts were detected in a further three regions (revealing evidence for upward shifts in 10 of 11 regions). Together, our results show that mountain environments are becoming increasingly exposed to biological invasions, emphasizing the need to monitor and prevent potential biosecurity issues emerging in high-elevation ecosystems.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Essl F, et al. A conceptual framework for range-expanding species that track human-induced environmental change. BioScience. 2019;69:908–919. doi: 10.1093/biosci/biz101. - DOI
-
- Freeman BG, Lee-Yaw JA, Sunday JM, Hargreaves AL. Expanding, shifting and shrinking: the impact of global warming on species’ elevational distributions. Glob. Ecol. Biogeogr. 2018;27:1268–1276. doi: 10.1111/geb.12774. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
