

Regulatory mechanisms of the dynein-2 motility by post-translational modification revealed by MD simulation
- PMID: 36702893
- PMCID: PMC9879972
- DOI: 10.1038/s41598-023-28026-z
Regulatory mechanisms of the dynein-2 motility by post-translational modification revealed by MD simulation
Abstract
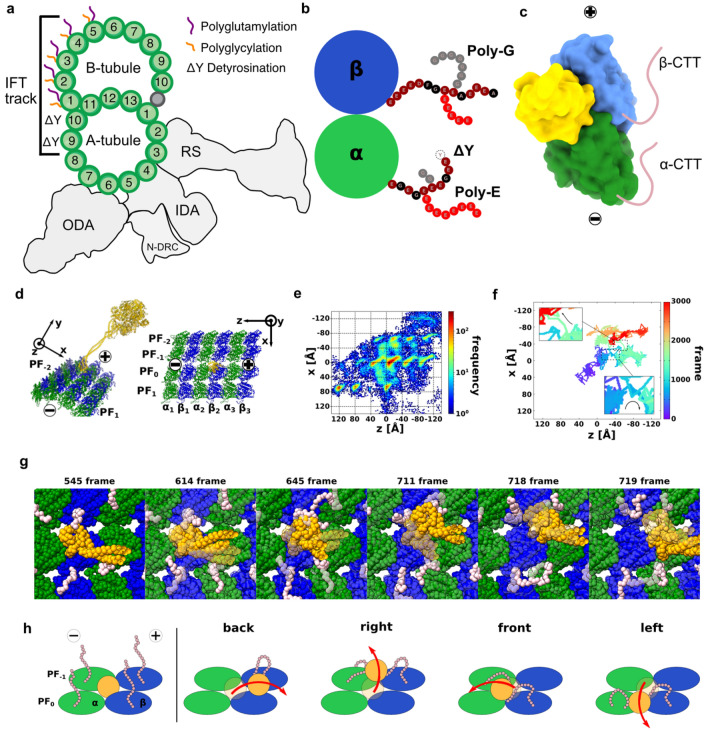
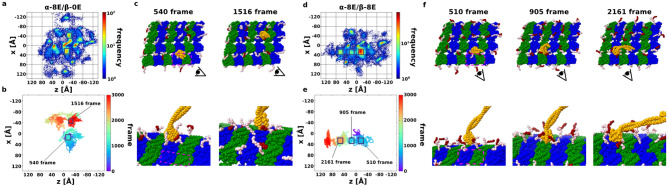
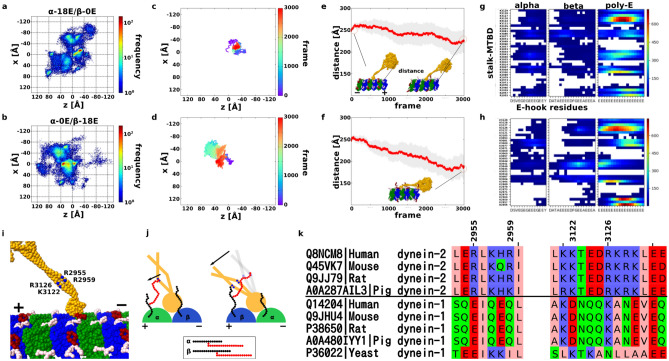
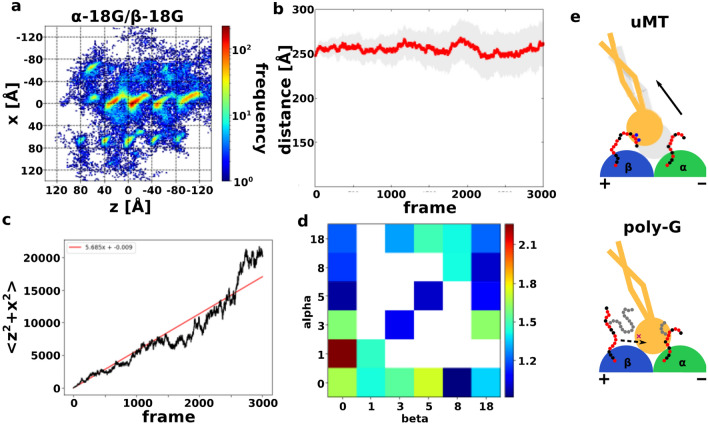
Intraflagellar transport for ciliary assembly and maintenance is driven by dynein and kinesins specific to the cilia. It has been shown that anterograde and retrograde transports run on different regions of the doublet microtubule, i.e., separate train tracks. However, little is known about the regulatory mechanism of this selective process. Since the doublet microtubule is known to display specific post-translational modifications of tubulins, i.e., "tubulin code", for molecular motor regulations, we investigated the motility of ciliary specific dynein-2 under different post-translational modification by coarse-grained molecular dynamics. Our setup allows us to simulate the landing behaviors of dynein-2 on un-modified, detyrosinated, poly-glutamylated and poly-glycylated microtubules in silico. Our study revealed that poly-glutamylation can play an inhibitory effect on dynein-2 motility. Our result indicates that poly-glutamylation of the B-tubule of the doublet microtubule can be used as an efficient means to target retrograde intraflagellar transport onto the A-tubule.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Tubulin glycylation controls ciliary motility through modulation of outer-arm dyneins.Mol Biol Cell. 2024 Jul 1;35(7):ar90. doi: 10.1091/mbc.E24-04-0154. Epub 2024 May 17. Mol Biol Cell. 2024. PMID: 38758663 Free PMC article.
-
Differential modification of the C-terminal tails of different α-tubulins and their importance for microtubule function in vivo.Elife. 2023 Jun 22;12:e87125. doi: 10.7554/eLife.87125. Elife. 2023. PMID: 37345829 Free PMC article.
-
Tubulin glutamylation regulates ciliary motility by altering inner dynein arm activity.Curr Biol. 2010 Mar 9;20(5):435-40. doi: 10.1016/j.cub.2009.12.062. Curr Biol. 2010. PMID: 20189389 Free PMC article.
-
Posttranslational Modifications of Tubulin and Cilia.Cold Spring Harb Perspect Biol. 2017 Jun 1;9(6):a028159. doi: 10.1101/cshperspect.a028159. Cold Spring Harb Perspect Biol. 2017. PMID: 28003186 Free PMC article. Review.
-
Ciliary and flagellar structure and function--their regulations by posttranslational modifications of axonemal tubulin.Int Rev Cell Mol Biol. 2012;294:133-70. doi: 10.1016/B978-0-12-394305-7.00003-3. Int Rev Cell Mol Biol. 2012. PMID: 22364873 Review.
Cited by
-
Theoretical insights into rotary mechanism of MotAB in the bacterial flagellar motor.Biophys J. 2024 Oct 15;123(20):3587-3599. doi: 10.1016/j.bpj.2024.09.010. Epub 2024 Sep 11. Biophys J. 2024. PMID: 39262115
-
Comparative Proteomic Analysis Reveals Altered Ciliary Proteins in Sickle Cell Disease.J Proteome Res. 2025 Jun 6;24(6):2981-2995. doi: 10.1021/acs.jproteome.5c00168. Epub 2025 May 15. J Proteome Res. 2025. PMID: 40374167
References
-
- Janke C, Magiera MM. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 2020;21(6):307–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
