

Spatial transcriptomics for profiling the tropism of viral vectors in tissues
- PMID: 36702899
- PMCID: PMC10443732
- DOI: 10.1038/s41587-022-01648-w
Spatial transcriptomics for profiling the tropism of viral vectors in tissues
Abstract
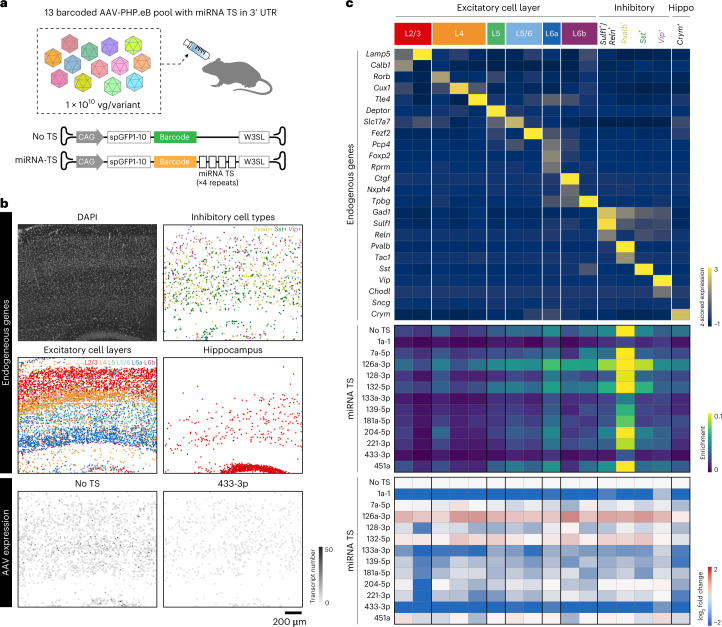
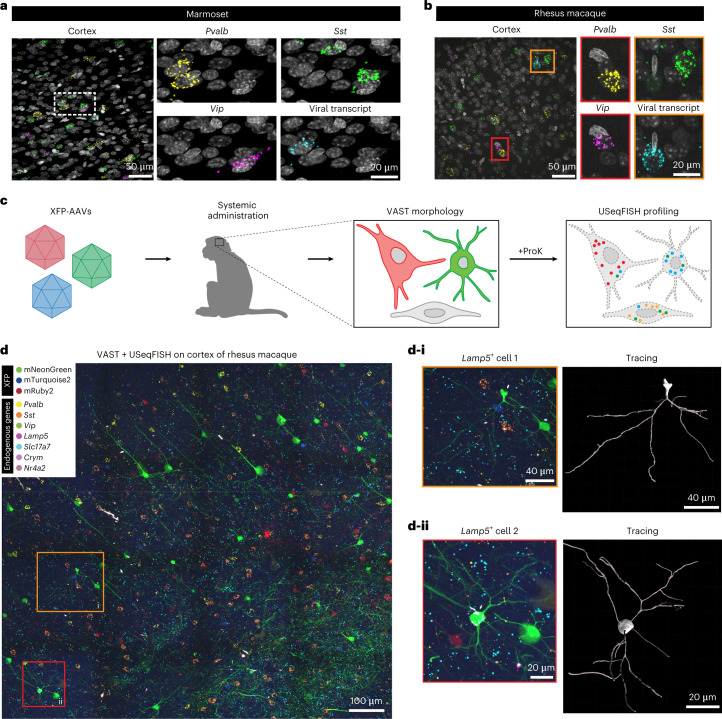
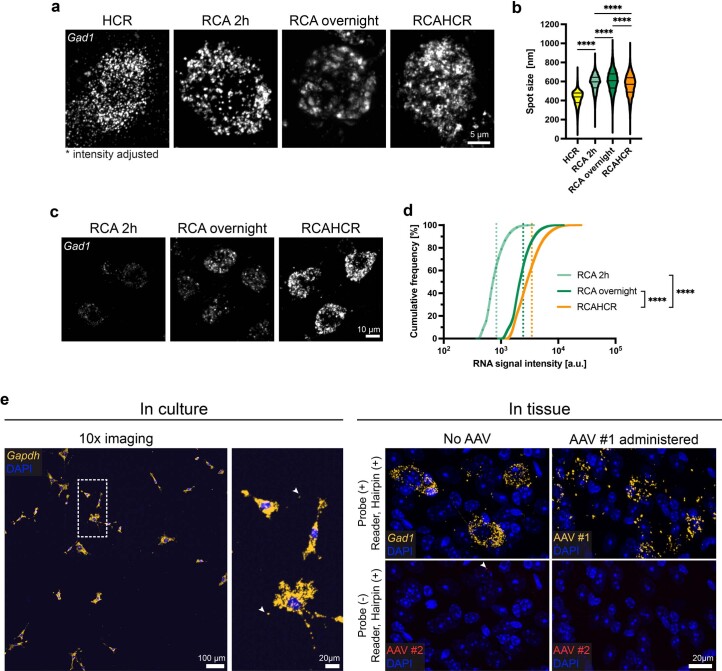
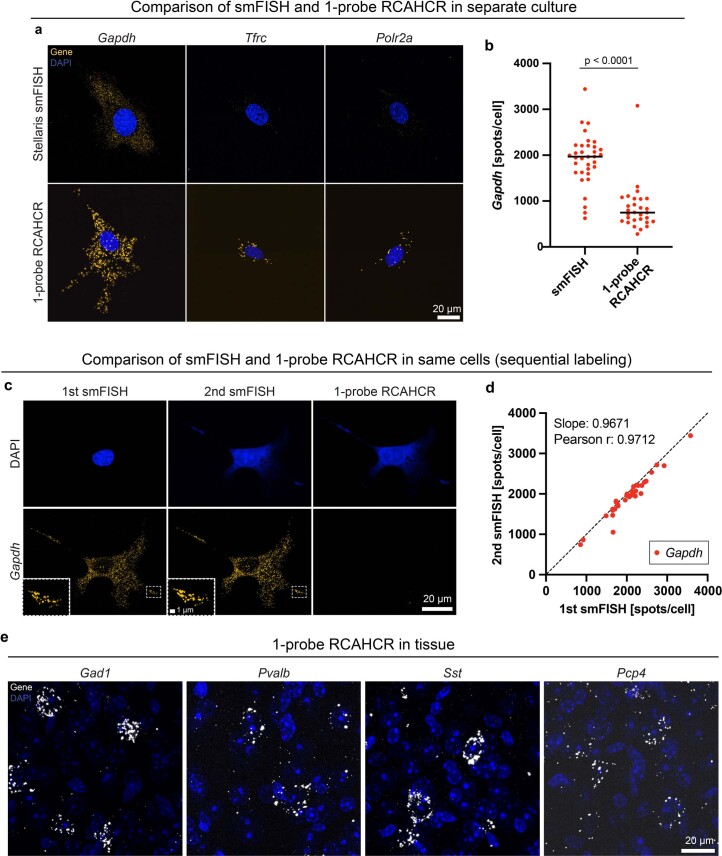
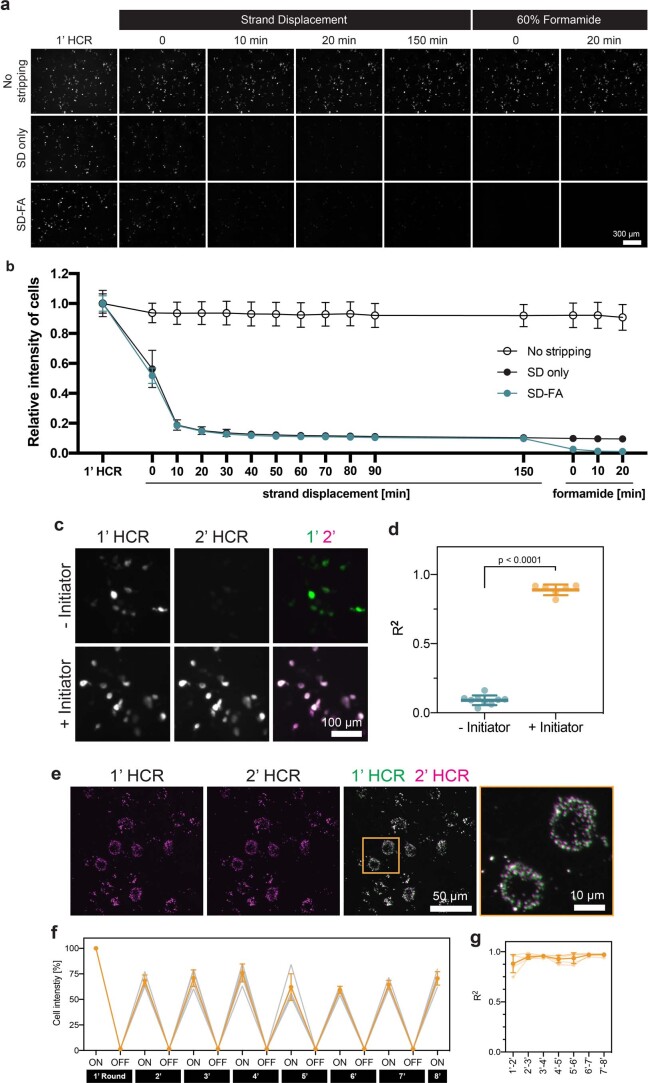
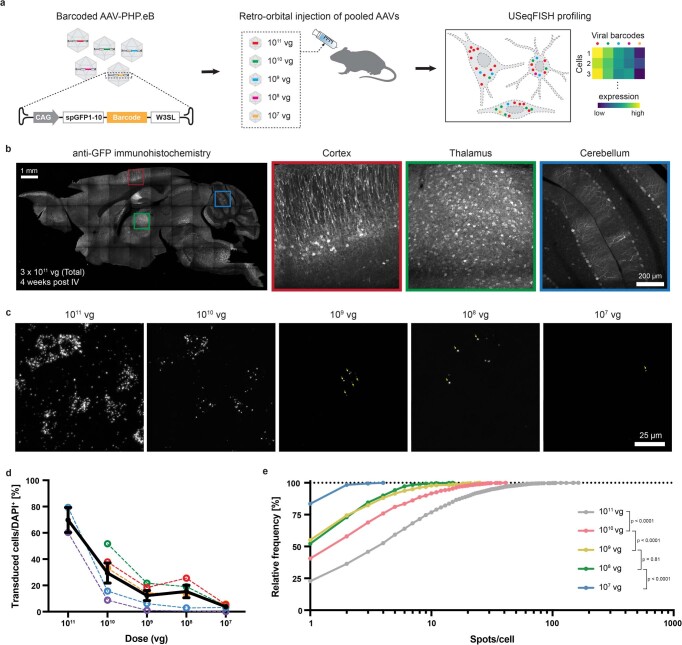
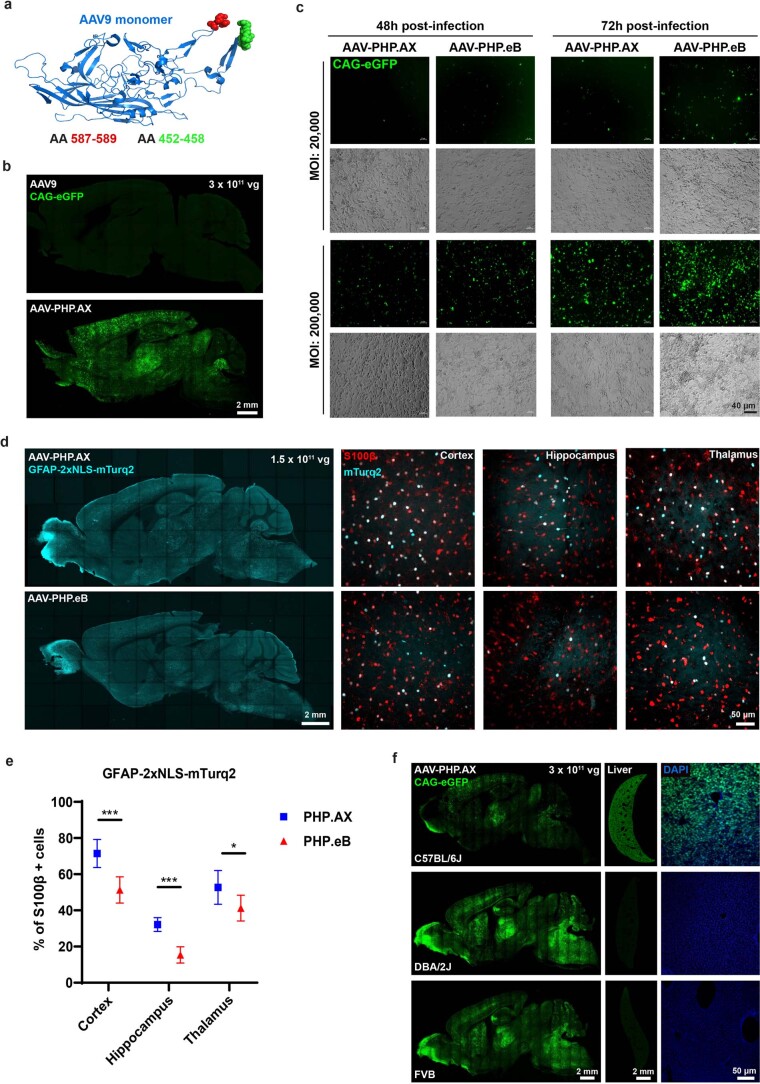
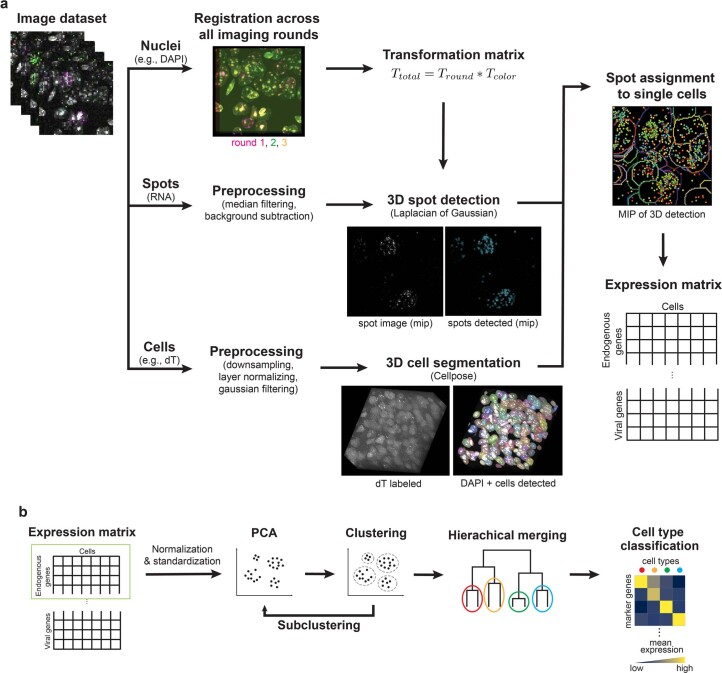
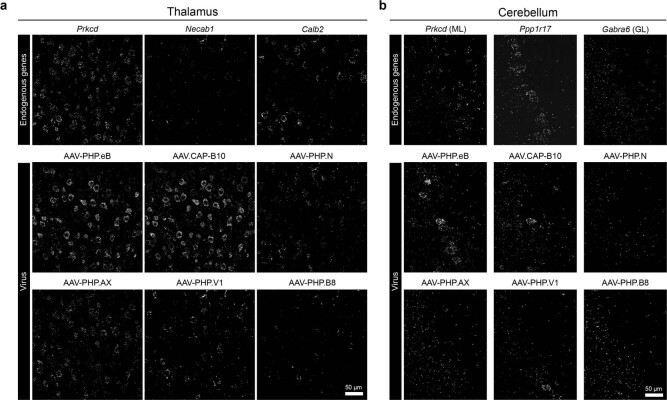
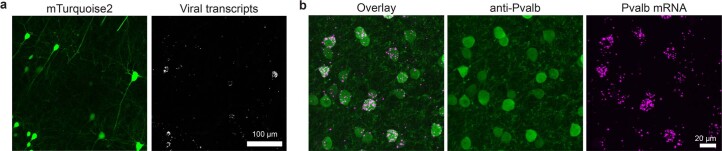
A barrier to advancing engineered adeno-associated viral vectors (AAVs) for precision access to cell subtypes is a lack of high-throughput, high-resolution assays to characterize in vivo transduction profiles. In this study, we developed an ultrasensitive, sequential fluorescence in situ hybridization (USeqFISH) method for spatial transcriptomic profiling of endogenous and viral RNA with a short barcode in intact tissue volumes by integrating hydrogel-based tissue clearing, enhanced signal amplification and multiplexing using sequential labeling. Using USeqFISH, we investigated the transduction and cell subtype tropisms across mouse brain regions of six systemic AAVs, including AAV-PHP.AX, a new variant that transduces robustly and efficiently across neurons and astrocytes. Here we reveal distinct cell subtype biases of each AAV variant, including a bias of AAV-PHP.N toward excitatory neurons. USeqFISH also enables profiling of pooled regulatory cargos, as we show for a 13-variant pool of microRNA target sites in AAV genomes. Lastly, we demonstrate potential applications of USeqFISH for in situ AAV profiling and multimodal single-cell analysis in non-human primates.
© 2023. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
V.G. is a co-founder and board member of Capsida Biotherapeutics, a fully integrated AAV engineering and gene therapy company. The remaining authors declare no competing interests.
Figures
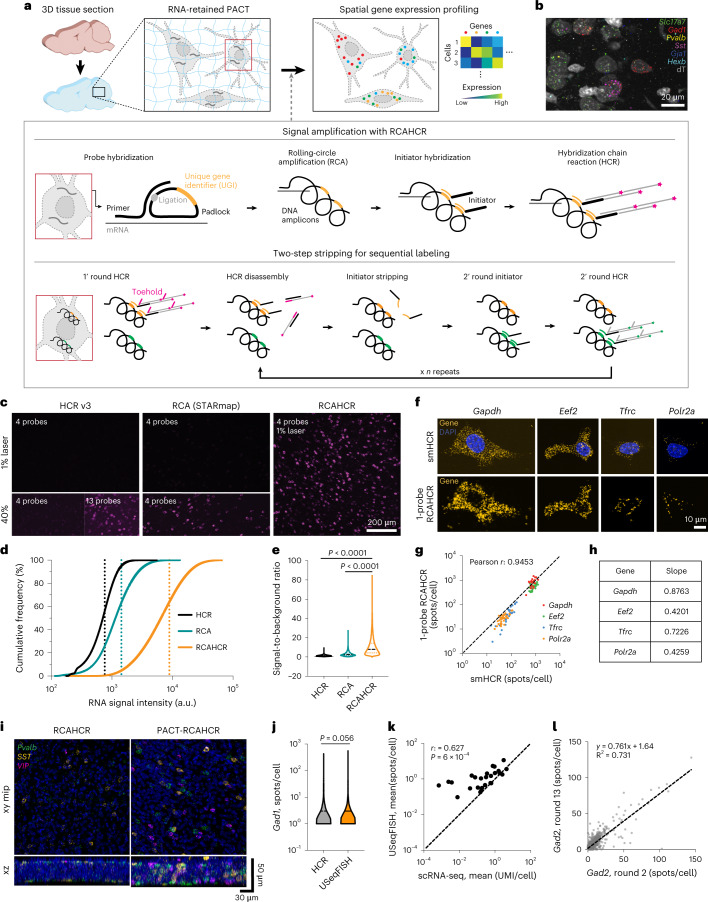
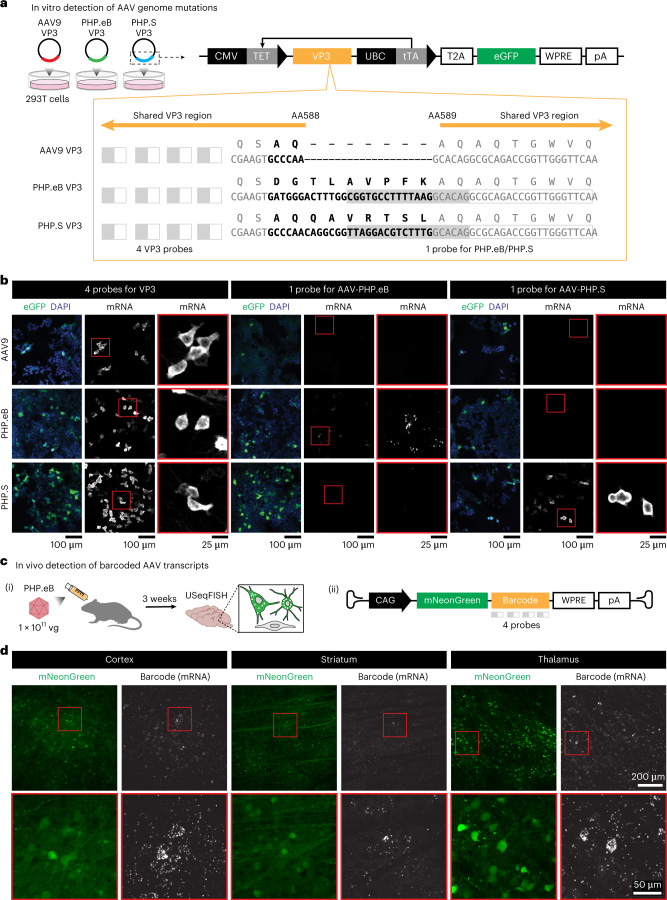
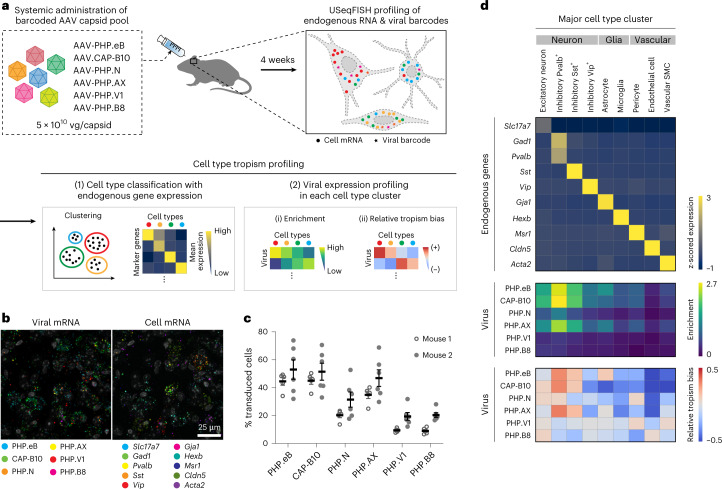
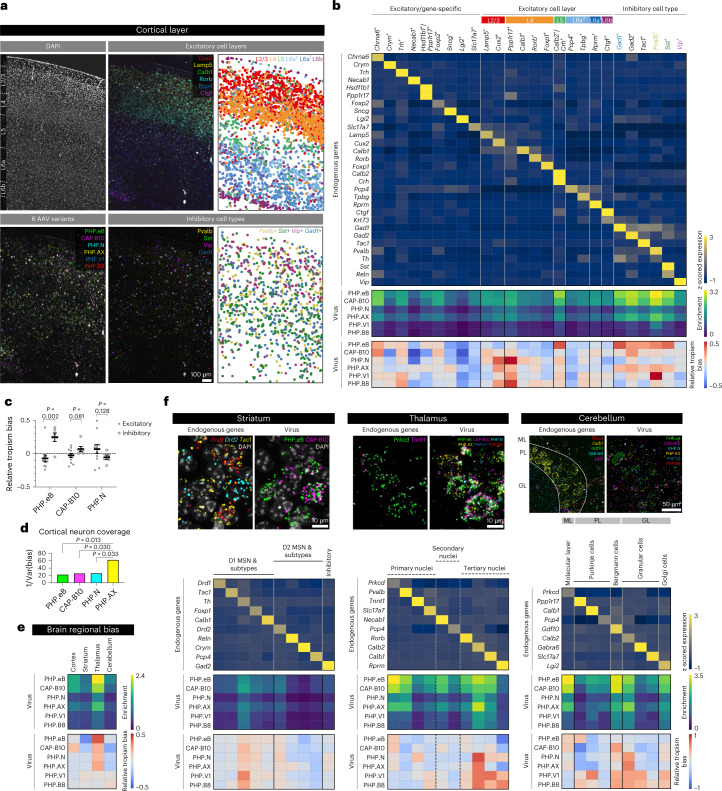

Comment in
-
Spatial tropism profiling of AAV vectors by ultrasensitive sequential FISH in tissue.Nat Biotechnol. 2023 Sep;41(9):1206-1207. doi: 10.1038/s41587-023-01681-3. Nat Biotechnol. 2023. PMID: 36717693 Free PMC article.
-
Profiling AAV tropism.Nat Rev Drug Discov. 2023 Mar;22(3):183. doi: 10.1038/d41573-023-00025-9. Nat Rev Drug Discov. 2023. PMID: 36755157 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
