

SUMOylation-mediated PSME3-20 S proteasomal degradation of transcription factor CP2c is crucial for cell cycle progression
- PMID: 36706181
- PMCID: PMC9882985
- DOI: 10.1126/sciadv.add4969
SUMOylation-mediated PSME3-20 S proteasomal degradation of transcription factor CP2c is crucial for cell cycle progression
Abstract
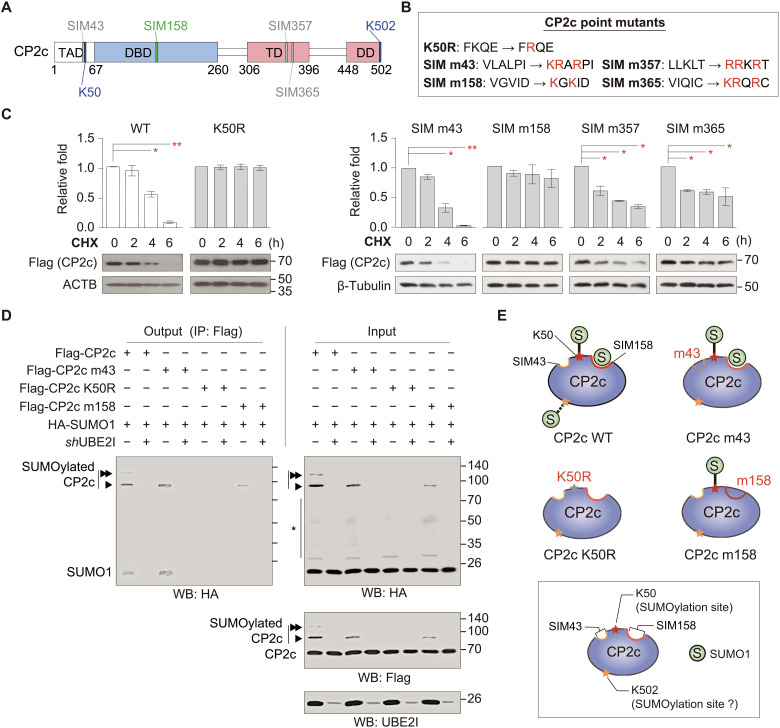
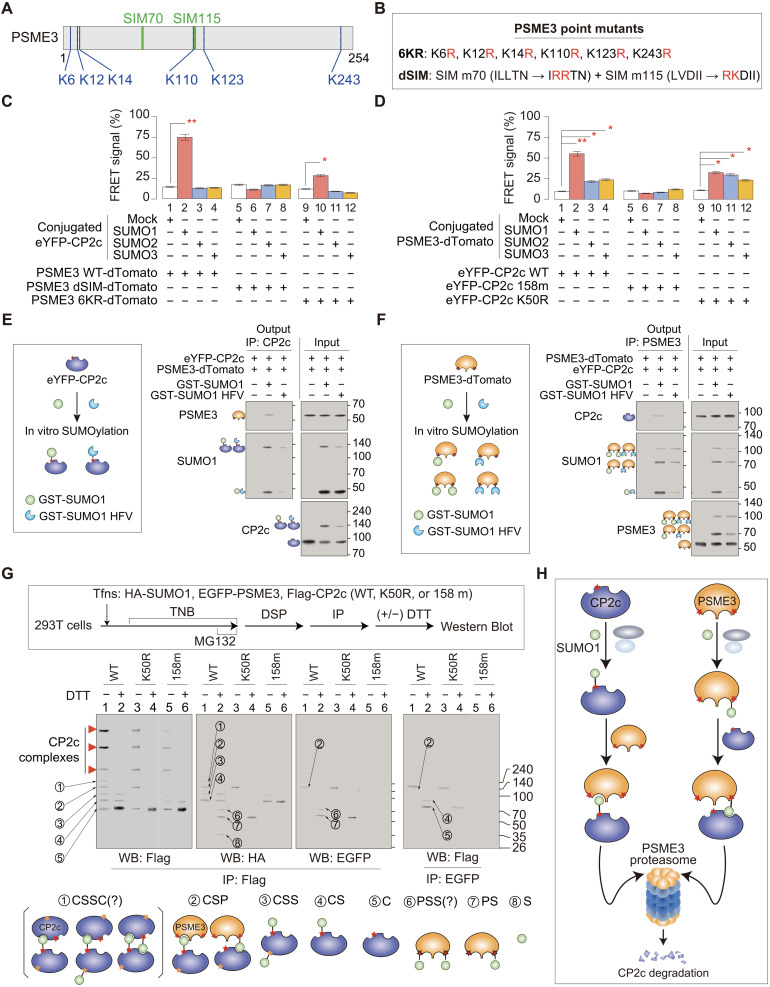
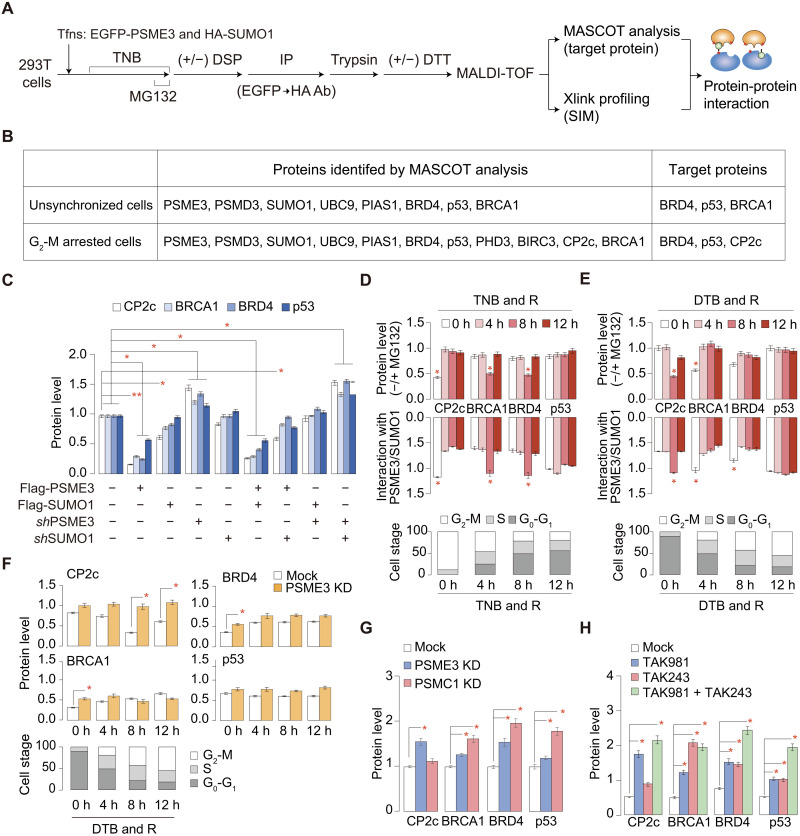
Transcription factor CP2c (also known as TFCP2, α-CP2, LSF, and LBP-1c) is involved in diverse ubiquitous and tissue/stage-specific cellular processes and in human malignancies such as cancer. Despite its importance, many fundamental regulatory mechanisms of CP2c are still unclear. Here, we uncover an unprecedented mechanism of CP2c degradation via a previously unidentified SUMO1/PSME3/20S proteasome pathway and its biological meaning. CP2c is SUMOylated in a SUMO1-dependent way, and SUMOylated CP2c is degraded through the ubiquitin-independent PSME3 (also known as REGγ or PA28)/20S proteasome system. SUMOylated PSME3 could also interact with CP2c to degrade CP2c via the 20S proteasomal pathway. Moreover, precisely timed degradation of CP2c via the SUMO1/PSME3/20S proteasome axis is required for accurate progression of the cell cycle. Therefore, we reveal a unique SUMO1-mediated uncanonical 20S proteasome degradation mechanism via the SUMO1/PSME3 axis involving mutual SUMO-SIM interaction of CP2c and PSME3, providing previously unidentified mechanistic insights into the roles of dynamic degradation of CP2c in cell cycle progression.
Figures



Similar articles
-
Structural and Functional Insights into CP2c Transcription Factor Complexes.Int J Mol Sci. 2022 Jun 7;23(12):6369. doi: 10.3390/ijms23126369. Int J Mol Sci. 2022. PMID: 35742810 Free PMC article.
-
Regulation of REGγ cellular distribution and function by SUMO modification.Cell Res. 2011 May;21(5):807-16. doi: 10.1038/cr.2011.57. Epub 2011 Mar 29. Cell Res. 2011. PMID: 21445096 Free PMC article.
-
Requirement of PML SUMO interacting motif for RNF4- or arsenic trioxide-induced degradation of nuclear PML isoforms.PLoS One. 2012;7(9):e44949. doi: 10.1371/journal.pone.0044949. Epub 2012 Sep 18. PLoS One. 2012. PMID: 23028697 Free PMC article.
-
Regulating the 20S proteasome ubiquitin-independent degradation pathway.Biomolecules. 2014 Sep 23;4(3):862-84. doi: 10.3390/biom4030862. Biomolecules. 2014. PMID: 25250704 Free PMC article. Review.
-
Regulation of Life & Death by REGγ.Cells. 2022 Jul 23;11(15):2281. doi: 10.3390/cells11152281. Cells. 2022. PMID: 35892577 Free PMC article. Review.
Cited by
-
SUMOylation of RALY promotes vasculogenic mimicry in glioma cells via the FOXD1/DKK1 pathway.Cell Biol Toxicol. 2023 Dec;39(6):3323-3340. doi: 10.1007/s10565-023-09836-3. Epub 2023 Oct 31. Cell Biol Toxicol. 2023. PMID: 37906341 Free PMC article.
-
Distinctive tumorigenic significance and innovative oncology targets of SUMOylation.Theranostics. 2024 May 19;14(8):3127-3149. doi: 10.7150/thno.97162. eCollection 2024. Theranostics. 2024. PMID: 38855173 Free PMC article. Review.
-
PQBP3 prevents senescence by suppressing PSME3-mediated proteasomal Lamin B1 degradation.EMBO J. 2024 Sep;43(18):3968-3999. doi: 10.1038/s44318-024-00192-4. Epub 2024 Aug 5. EMBO J. 2024. PMID: 39103492 Free PMC article.
-
Mevalonate pathway inhibition reduces bladder cancer metastasis by modulating RhoB protein stability and integrin β1 localization.Commun Biol. 2024 Nov 9;7(1):1476. doi: 10.1038/s42003-024-07067-8. Commun Biol. 2024. PMID: 39521858 Free PMC article.
-
O-GlcNAcylation: the sweet side of epigenetics.Epigenetics Chromatin. 2023 Dec 14;16(1):49. doi: 10.1186/s13072-023-00523-5. Epigenetics Chromatin. 2023. PMID: 38093337 Free PMC article. Review.
References
-
- Rodda S., Sharma S., Scherer M., Chapman G., Rathjen P., CRTR-1, a developmentally regulated transcriptional repressor related to the CP2 family of transcription factors. J. Biol. Chem. 276, 3324–3332 (2001). - PubMed
-
- Wilanowski T., Tuckfield A., Cerruti L., O’Connell S., Saint R., Parekh V., Tao J., Cunningham J. M., Jane S. M., A highly conserved novel family of mammalian developmental transcription factors related to Drosophila grainyhead. Mech. Dev. 114, 37–50 (2002). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
