

Redox-dependent Igfbp2 signaling controls Brca1 DNA damage response to govern neural stem cell fate
- PMID: 36707536
- PMCID: PMC9883463
- DOI: 10.1038/s41467-023-36174-z
Redox-dependent Igfbp2 signaling controls Brca1 DNA damage response to govern neural stem cell fate
Abstract
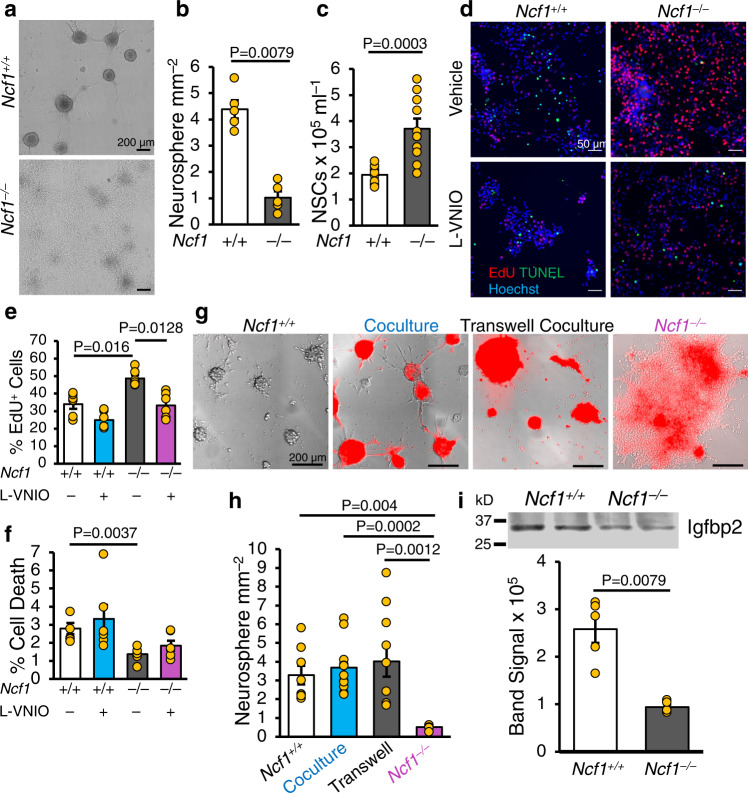
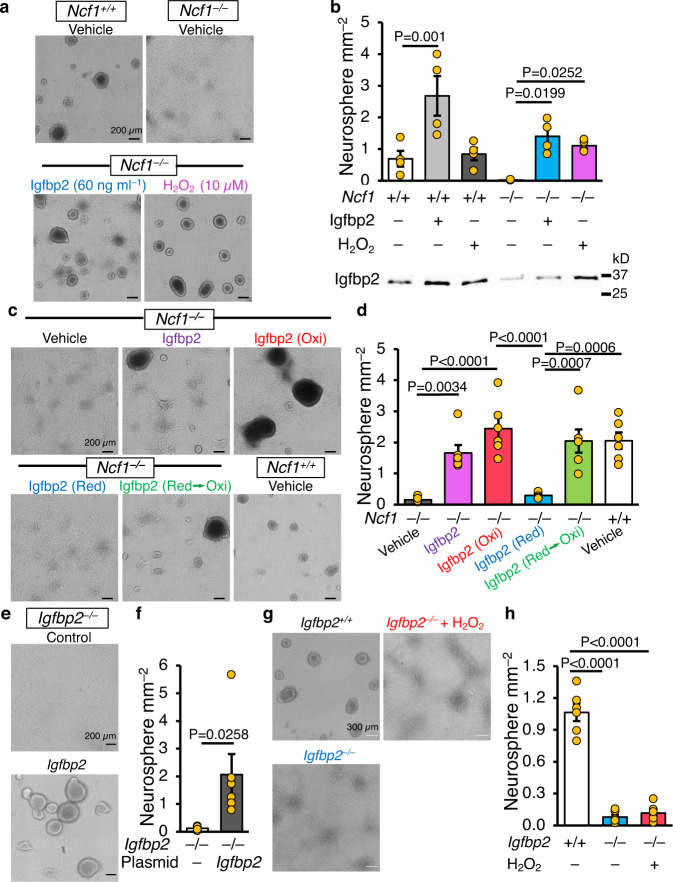
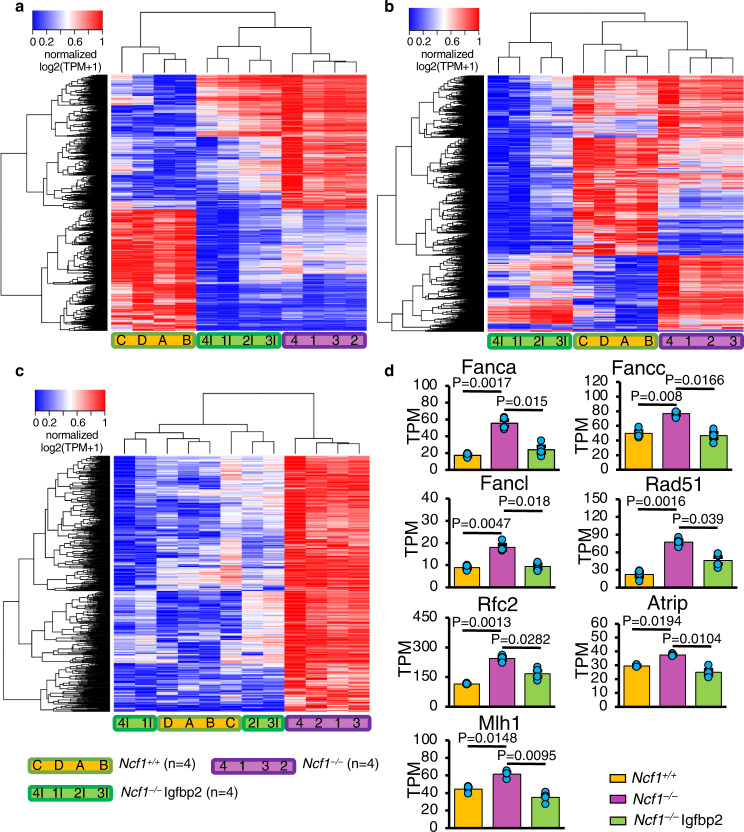
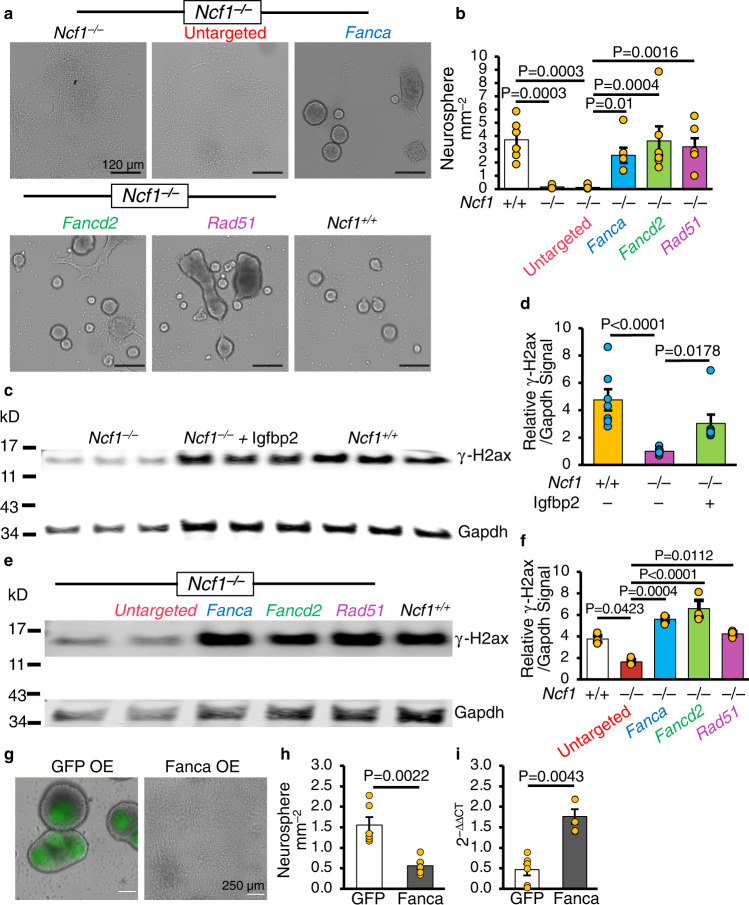
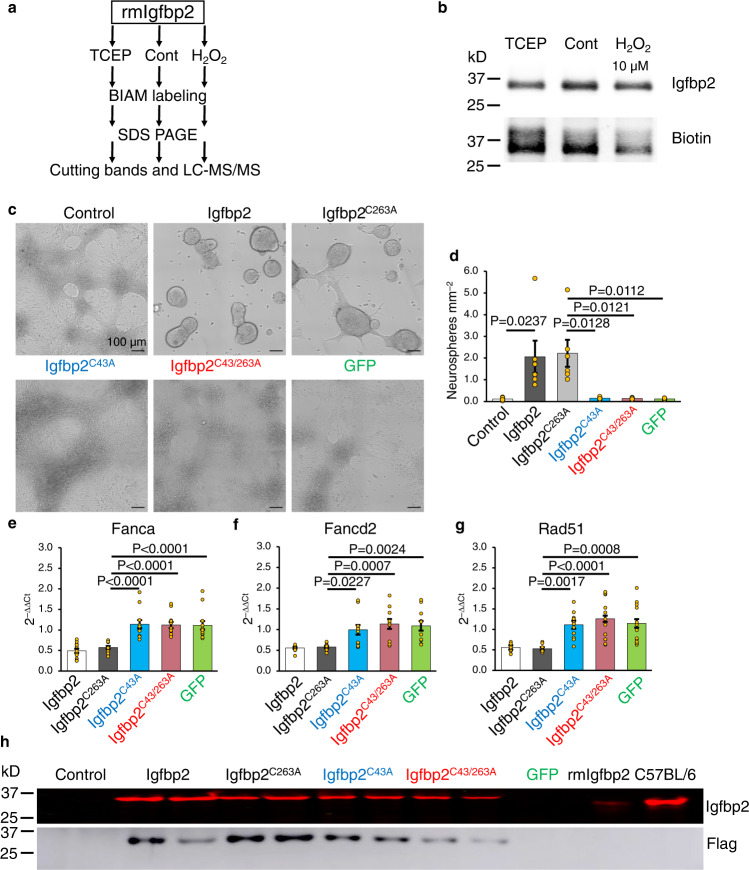
Neural stem cell (NSC) maintenance and functions are regulated by reactive oxygen species (ROS). However, the mechanisms by which ROS control NSC behavior remain unclear. Here we report that ROS-dependent Igfbp2 signaling controls DNA repair pathways which balance NSC self-renewal and lineage commitment. Ncf1 or Igfbp2 deficiency constrains NSCs to a self-renewing state and prevents neurosphere formation. Ncf1-dependent oxidation of Igfbp2 promotes neurogenesis by NSCs in vitro and in vivo while repressing Brca1 DNA damage response genes and inducing DNA double-strand breaks (DDSBs). By contrast, Ncf1-/- and Igfbp2-/- NSCs favor the formation of oligodendrocytes in vitro and in vivo. Notably, transient repression of Brca1 DNA repair pathway genes induces DDSBs and is sufficient to rescue the ability of Ncf1-/- and Igfbp2-/- NSCs to lineage-commit to form neurospheres and neurons. NSC lineage commitment is dependent on the oxidizable cysteine-43 residue of Igfbp2. Our study highlights the role of DNA damage/repair in orchestrating NSC fate decisions downstream of redox-regulated Igfbp2.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
IGFBP2 promotes neural stem cell maintenance and proliferation differentially associated with glioblastoma subtypes.Brain Res. 2019 Feb 1;1704:174-186. doi: 10.1016/j.brainres.2018.10.018. Epub 2018 Oct 19. Brain Res. 2019. PMID: 30347220
-
53BP1 regulates the self-renewal ability of neural stem/progenitor cells through modulating mitochondrial homeostasis.Biochem Biophys Res Commun. 2024 Nov 19;734:150776. doi: 10.1016/j.bbrc.2024.150776. Epub 2024 Oct 2. Biochem Biophys Res Commun. 2024. PMID: 39368367
-
Amyloid β Peptide Compromises Neural Stem Cell Fate by Irreversibly Disturbing Mitochondrial Oxidative State and Blocking Mitochondrial Biogenesis and Dynamics.Mol Neurobiol. 2019 Jun;56(6):3922-3936. doi: 10.1007/s12035-018-1342-z. Epub 2018 Sep 18. Mol Neurobiol. 2019. PMID: 30225776
-
Neural Stem Cells and Nutrients: Poised Between Quiescence and Exhaustion.Trends Endocrinol Metab. 2016 Nov;27(11):756-769. doi: 10.1016/j.tem.2016.06.007. Epub 2016 Jul 4. Trends Endocrinol Metab. 2016. PMID: 27387597 Review.
-
Epigenetic plasticity and redox regulation of neural stem cell state and fate.Free Radic Biol Med. 2021 Jul;170:116-130. doi: 10.1016/j.freeradbiomed.2021.02.030. Epub 2021 Mar 6. Free Radic Biol Med. 2021. PMID: 33684459 Review.
Cited by
-
Knockdown of KDM5B Leads to DNA Damage and Cell Cycle Arrest in Granulosa Cells via MTF1.Curr Issues Mol Biol. 2023 Apr 7;45(4):3219-3237. doi: 10.3390/cimb45040210. Curr Issues Mol Biol. 2023. PMID: 37185734 Free PMC article.
-
Defective Neurogenesis in Lowe Syndrome is Caused by Mitochondria Loss and Cilia-related Sonic Hedgehog Defects.bioRxiv [Preprint]. 2024 Nov 1:2024.11.01.621496. doi: 10.1101/2024.11.01.621496. bioRxiv. 2024. PMID: 39553960 Free PMC article. Preprint.
-
IGFBP-2 and IGF-II: Key Components of the Neural Stem Cell Niche? Implications for Glioblastoma Pathogenesis.Int J Mol Sci. 2025 May 15;26(10):4749. doi: 10.3390/ijms26104749. Int J Mol Sci. 2025. PMID: 40429889 Free PMC article. Review.
-
DNA damage and repair: underlying mechanisms leading to microcephaly.Front Cell Dev Biol. 2023 Oct 10;11:1268565. doi: 10.3389/fcell.2023.1268565. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37881689 Free PMC article. Review.
-
The dual role of cellular senescence in human tumor progression and therapy.MedComm (2020). 2024 Aug 19;5(9):e695. doi: 10.1002/mco2.695. eCollection 2024 Sep. MedComm (2020). 2024. PMID: 39161800 Free PMC article. Review.
References
-
- Ito K, et al. Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature. 2004;431:997–1002. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
