

Chronic stress in solid tumor development: from mechanisms to interventions
- PMID: 36707854
- PMCID: PMC9883141
- DOI: 10.1186/s12929-023-00903-9
Chronic stress in solid tumor development: from mechanisms to interventions
Abstract
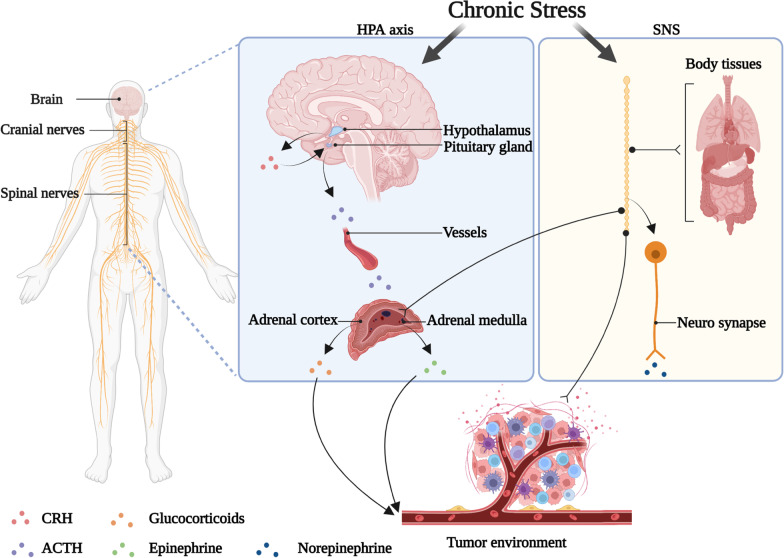
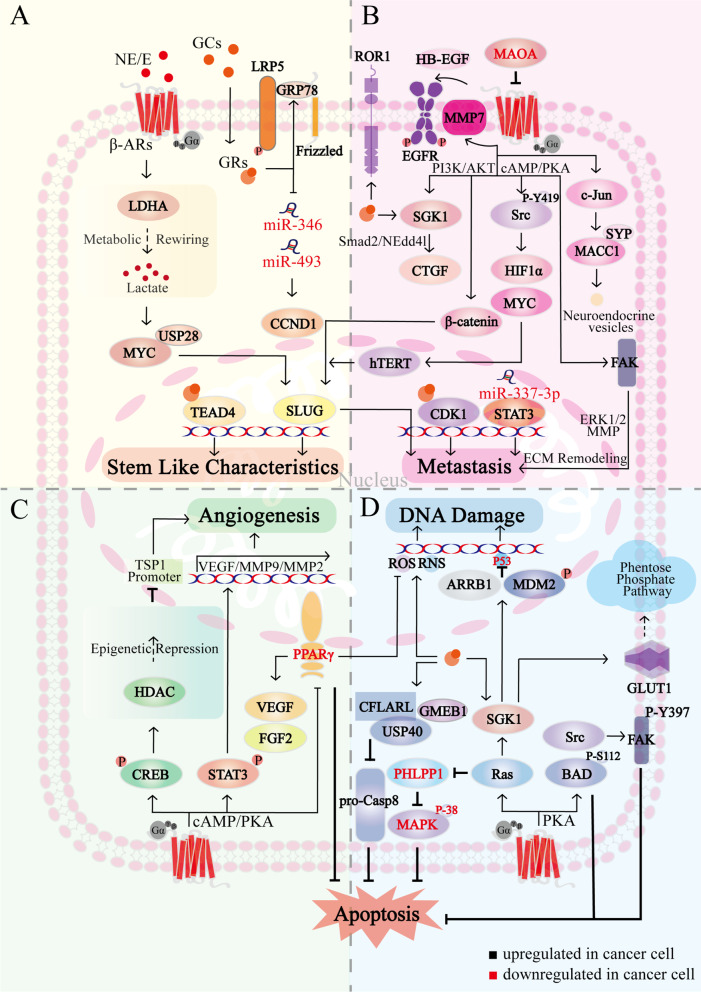
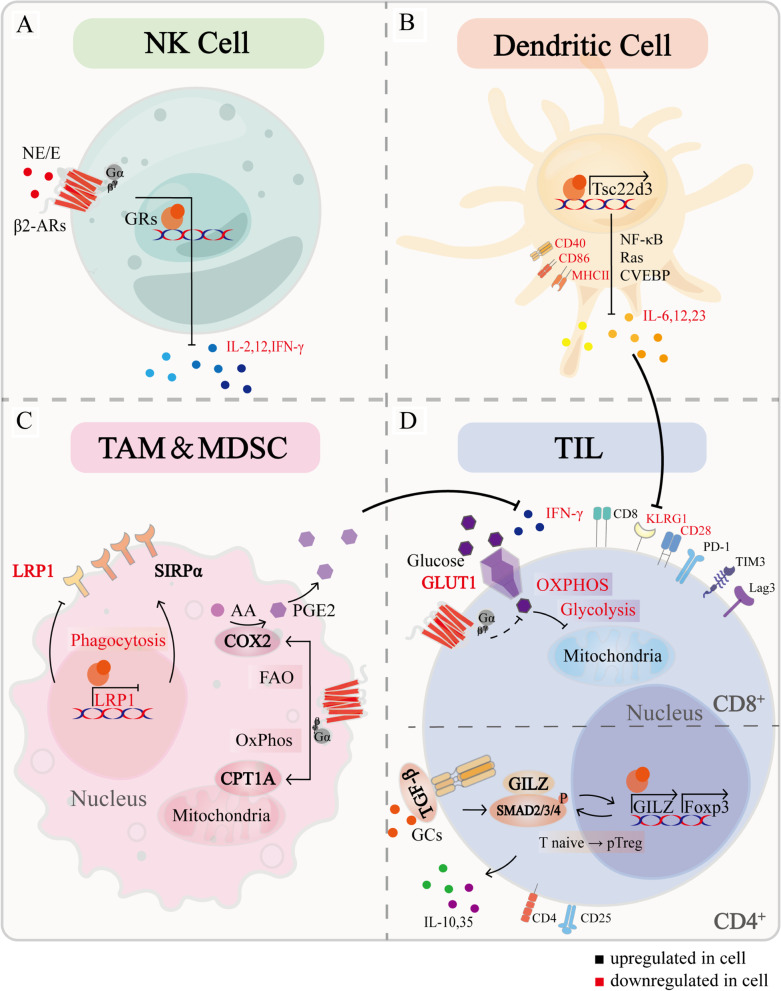
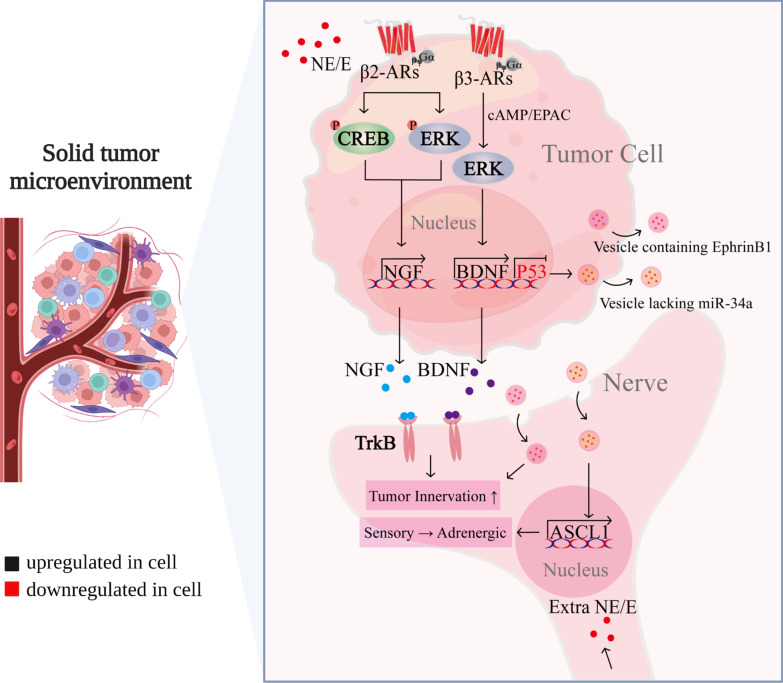
Chronic stress results in disturbances of body hormones through the neuroendocrine system. Cancer patients often experience recurrent anxiety and restlessness during disease progression and treatment, which aggravates disease progression and hinders treatment effects. Recent studies have shown that chronic stress-regulated neuroendocrine systems secret hormones to activate many signaling pathways related to tumor development in tumor cells. The activated neuroendocrine system acts not only on tumor cells but also modulates the survival and metabolic changes of surrounding non-cancerous cells. Current clinical evidences also suggest that chronic stress affects the outcome of cancer treatment. However, in clinic, there is lack of effective treatment for chronic stress in cancer patients. In this review, we discuss the main mechanisms by which chronic stress regulates the tumor microenvironment, including functional regulation of tumor cells by stress hormones (stem cell-like properties, metastasis, angiogenesis, DNA damage accumulation, and apoptotic resistance), metabolic reprogramming and immune escape, and peritumor neuromodulation. Based on the current clinical treatment framework for cancer and chronic stress, we also summarize pharmacological and non-pharmacological therapeutic approaches to provide some directions for cancer therapy.
Keywords: Cancer treatment; Chronic stress; Tumor development; Tumor microenvironment.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Yuan K, Zheng YB, Wang YJ, Sun YK, Gong YM, Huang YT, Chen X, Liu XX, Zhong Y, Su SZ, et al. A systematic review and meta-analysis on prevalence of and risk factors associated with depression, anxiety and insomnia in infectious diseases, including COVID-19: a call to action. Mol Psychiatry. 2022 doi: 10.1038/s41380-022-01638-z. - DOI - PMC - PubMed
-
- Russell G, Lightman S. The human stress response. Nat Rev Endocrinol. 2019;15(9):525–534. - PubMed
-
- Cui B, Peng F, Lu J, He B, Su Q, Luo H, Deng Z, Jiang T, Su K, Huang Y, et al. Cancer and stress: NextGen strategies. Brain Behav Immun. 2021;93:368–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 81972479/National Natural Science Foundation of China
- U2004118/National Natural Science Foundation of China
- 82072899/National Natural Science Foundation of China
- 2019A1515011100/Natural Science Foundation of Guangdong Province
- 2021A1515012576/Natural Science Foundation of Guangdong Province
- 202300410359/Henan Natural Science Foundation
- SBGJ2020002081/Henan Medical Research Program
- 202005/Guangzhou High-Level Clinical Key Specialty Construction Project: Clinical Key Specialty Construction Project of Guangzhou Medical University
- 2021KTSCX026/the Innovation Project of Universities in Guangdong Province
LinkOut - more resources
Full Text Sources
Medical
