

Lack of a protective effect of the Tmem106b "protective SNP" in the Grn knockout mouse model for frontotemporal lobar degeneration
- PMID: 36707901
- PMCID: PMC9881268
- DOI: 10.1186/s40478-023-01510-3
Lack of a protective effect of the Tmem106b "protective SNP" in the Grn knockout mouse model for frontotemporal lobar degeneration
Abstract
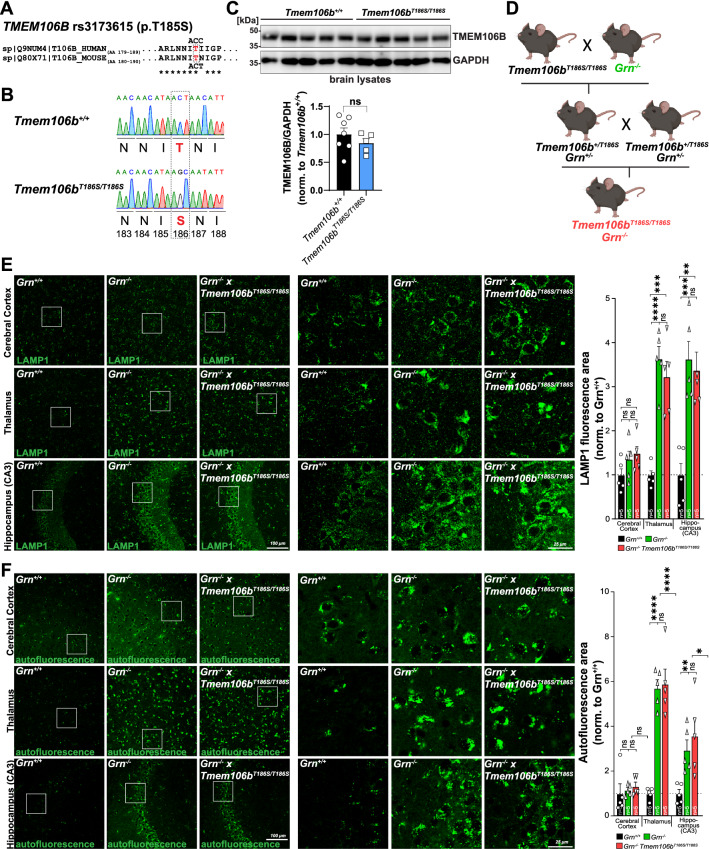
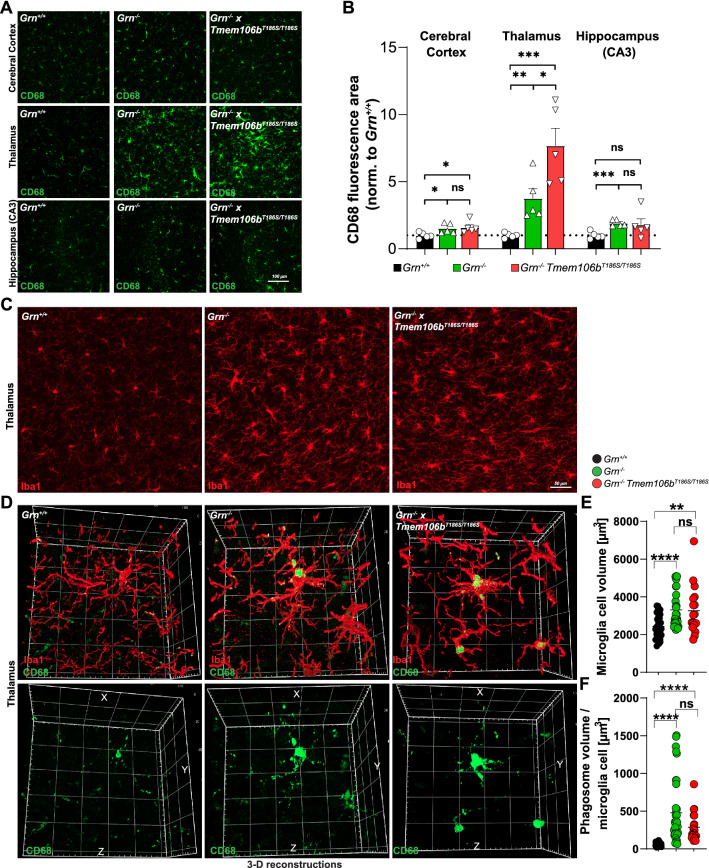
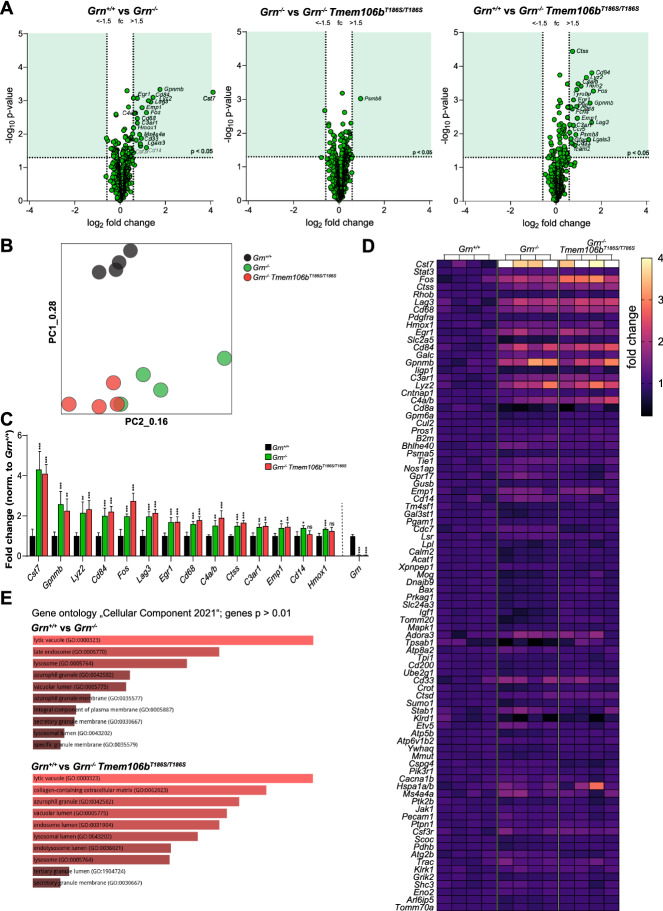
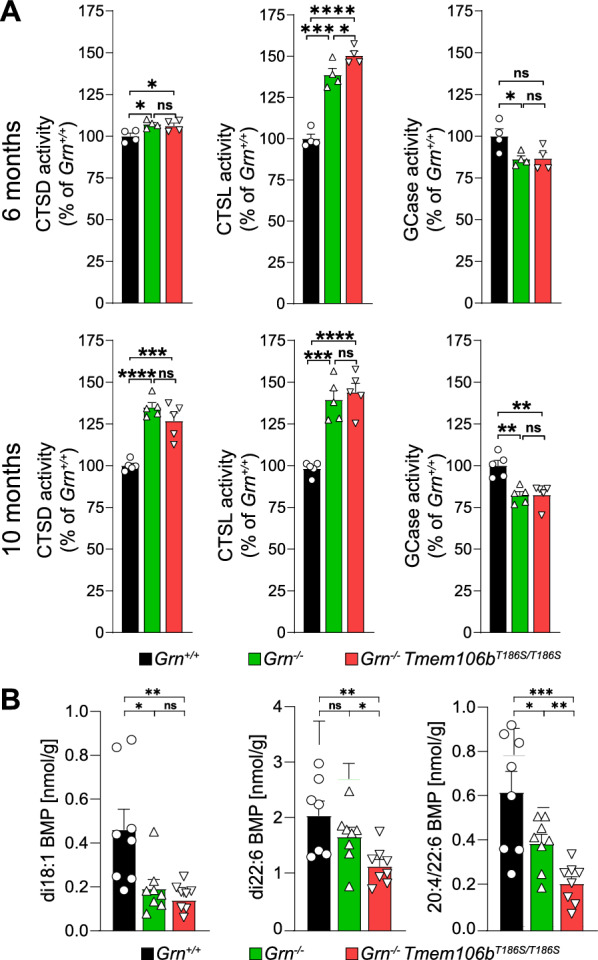
Genetic variants in TMEM106B are a common risk factor for frontotemporal lobar degeneration and the most important modifier of disease risk in patients with progranulin (GRN) mutations (FTLD-GRN). TMEM106B is encoding a lysosomal transmembrane protein of unknown molecular function. How it mediates its disease-modifying function remains enigmatic. Several TMEM106B single nucleotide polymorphisms (SNPs) are significantly associated with disease risk in FTLD-GRN carriers, of which all except one are within intronic sequences of TMEM106B. Of note, the non-coding SNPs are in high linkage disequilibrium with the coding SNP rs3173615 located in exon six of TMEM106B, resulting in a threonine to serine change at amino acid 185 in the minor allele, which is protective in FTLD-GRN carriers. To investigate the functional consequences of this variant in vivo, we generated and characterized a knockin mouse model harboring the Tmem106bT186S variant. We analyzed the effect of this protective variant on FTLD pathology by crossing Tmem106bT186S mice with Grn-/- knockout mice, a model for GRN-mediated FTLD. We did not observe the amelioration of any of the investigated Grn-/- knockout phenotypes, including transcriptomic changes, lipid alterations, or microgliosis in Tmem106bT186S/T186S × Grn-/- mice, indicating that the Tmem106bT186S variant is not protective in the Grn-/- knockout mouse model. These data suggest that effects of the associated SNPs not directly linked to the amino acid exchange in TMEM106B are critical for the modifying effect.
Keywords: FTLD; Progranulin; Protective SNP; TMEM106B.
© 2023. The Author(s).
Conflict of interest statement
MD participated in advisory board meetings of Arkuda Therapeutics and received a speaker honorarium from Takeda.
Figures
References
-
- Ahmed Z, Sheng H, Xu YF, Lin WL, Innes AE, Gass J, Yu X, Wuertzer CA, Hou H, Chiba S, et al. Accelerated lipofuscinosis and ubiquitination in granulin knockout mice suggest a role for progranulin in successful aging. Am J Pathol. 2010;177:311–324. doi: 10.2353/ajpath.2010.090915. - DOI - PMC - PubMed
-
- Arrant AE, Roth JR, Boyle NR, Kashyap SN, Hoffmann MQ, Murchison CF, Ramos EM, Nana AL, Spina S, Grinberg LT, et al. Impaired beta-glucocerebrosidase activity and processing in frontotemporal dementia due to progranulin mutations. Acta Neuropathol Commun. 2019;7:218. doi: 10.1186/s40478-019-0872-6. - DOI - PMC - PubMed
-
- Bellenguez C, Kucukali F, Jansen IE, Kleineidam L, Moreno-Grau S, Amin N, Naj AC, Campos-Martin R, Grenier-Boley B, Andrade V, et al. New insights into the genetic etiology of Alzheimer's disease and related dementias. Nat Genet. 2022;54:412–436. doi: 10.1038/s41588-022-01024-z. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
