

Specialized cells that sense tissue mechanics to regulate Drosophila morphogenesis
- PMID: 36708706
- PMCID: PMC11345685
- DOI: 10.1016/j.devcel.2023.01.004
Specialized cells that sense tissue mechanics to regulate Drosophila morphogenesis
Abstract
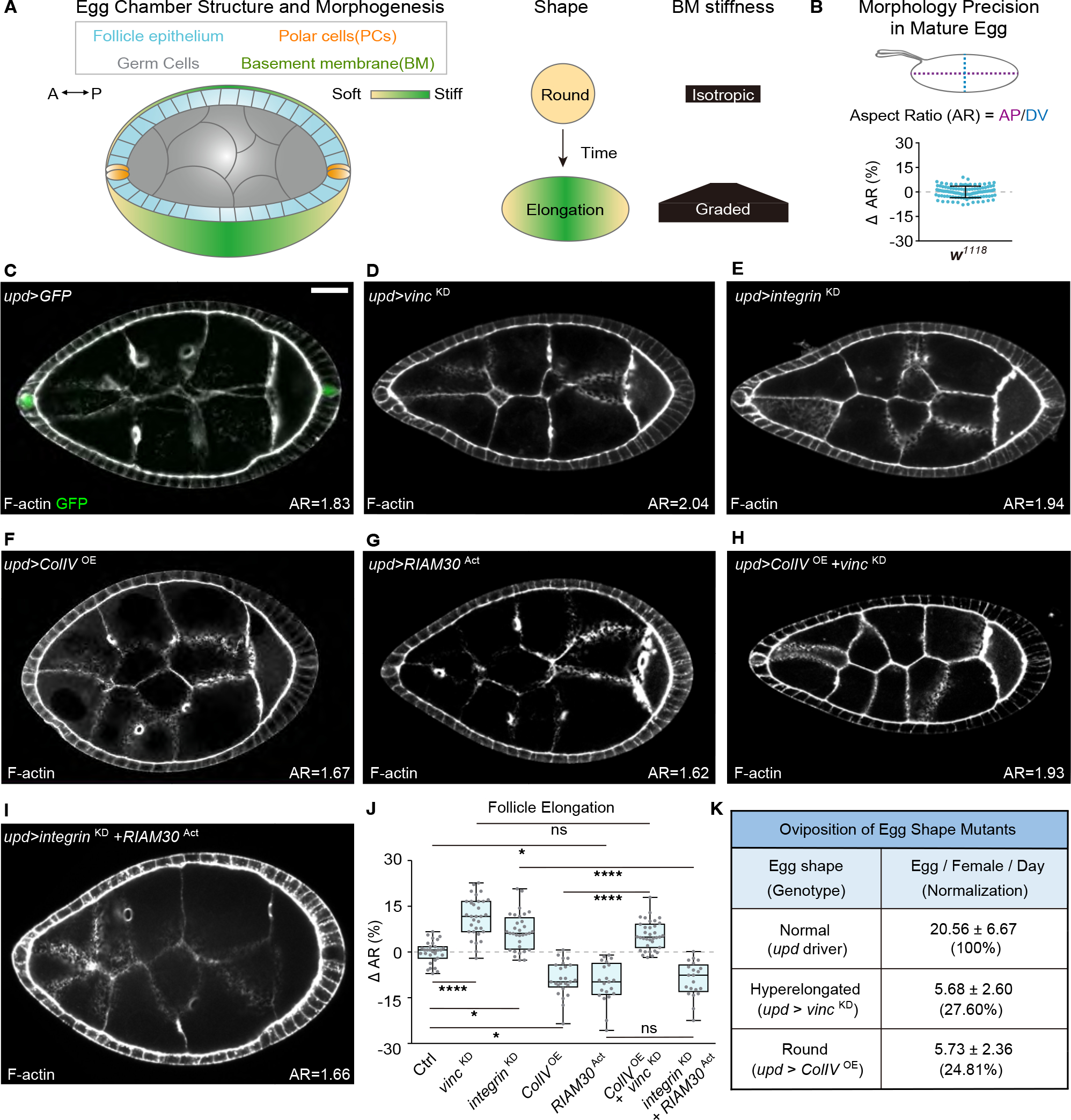
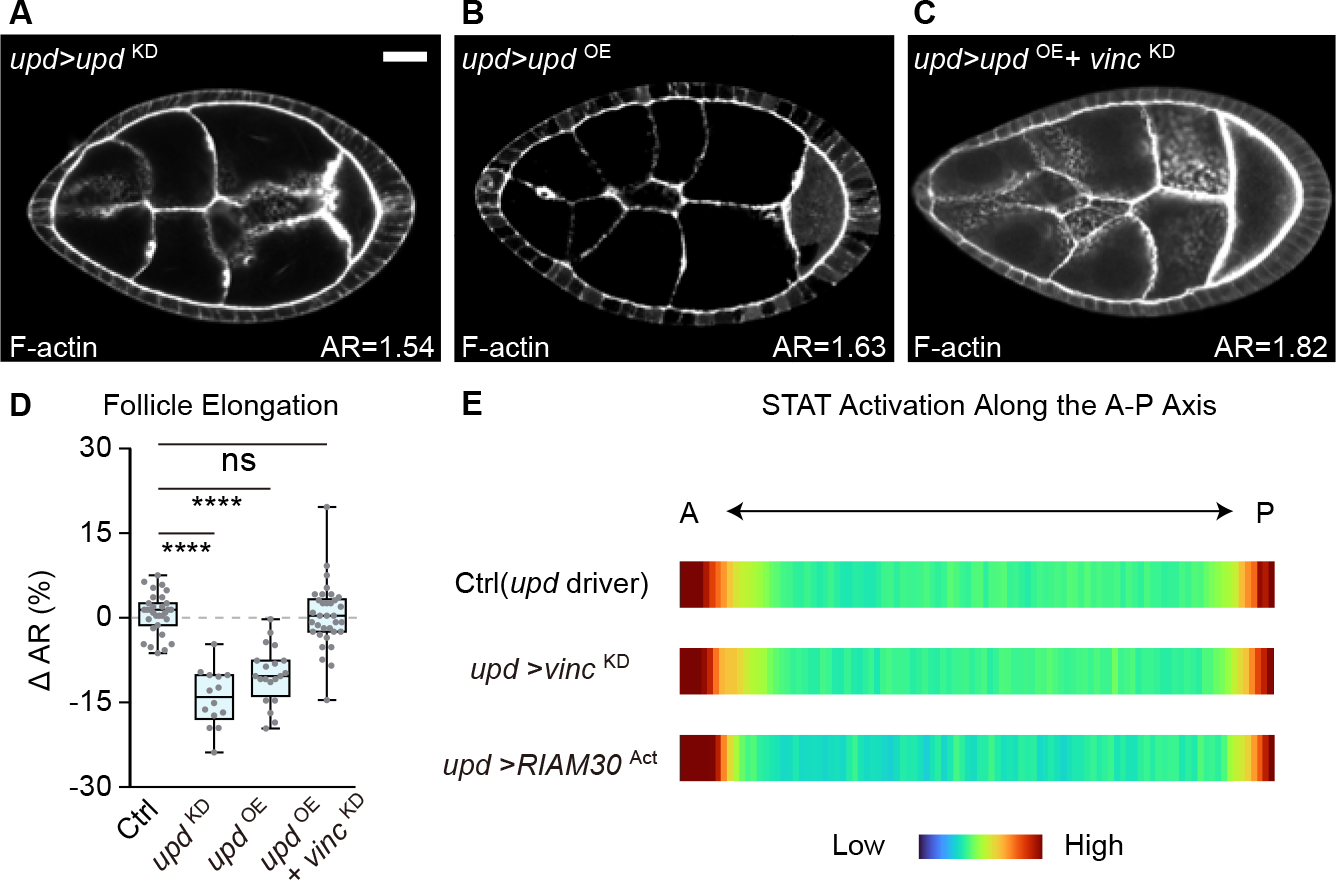
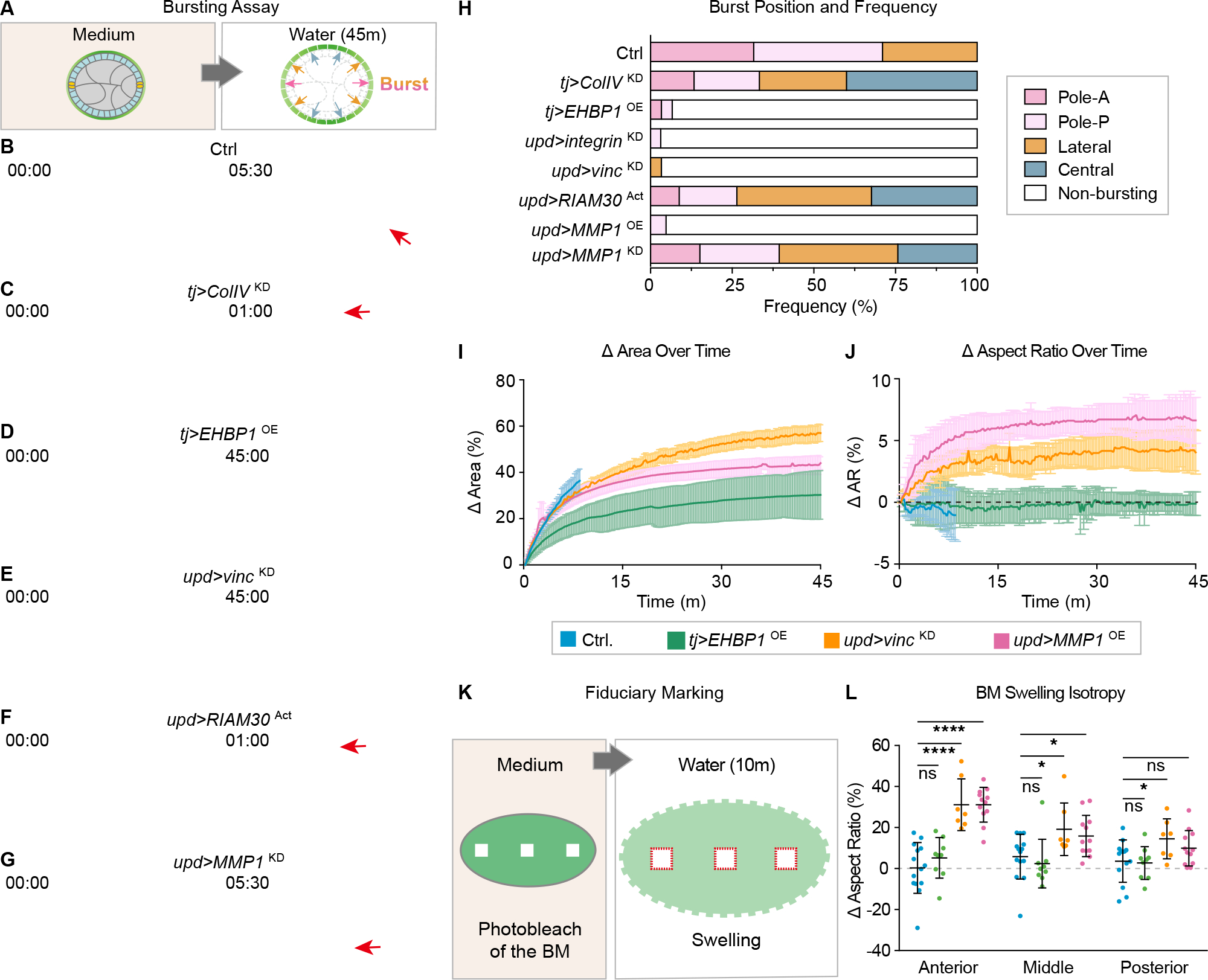
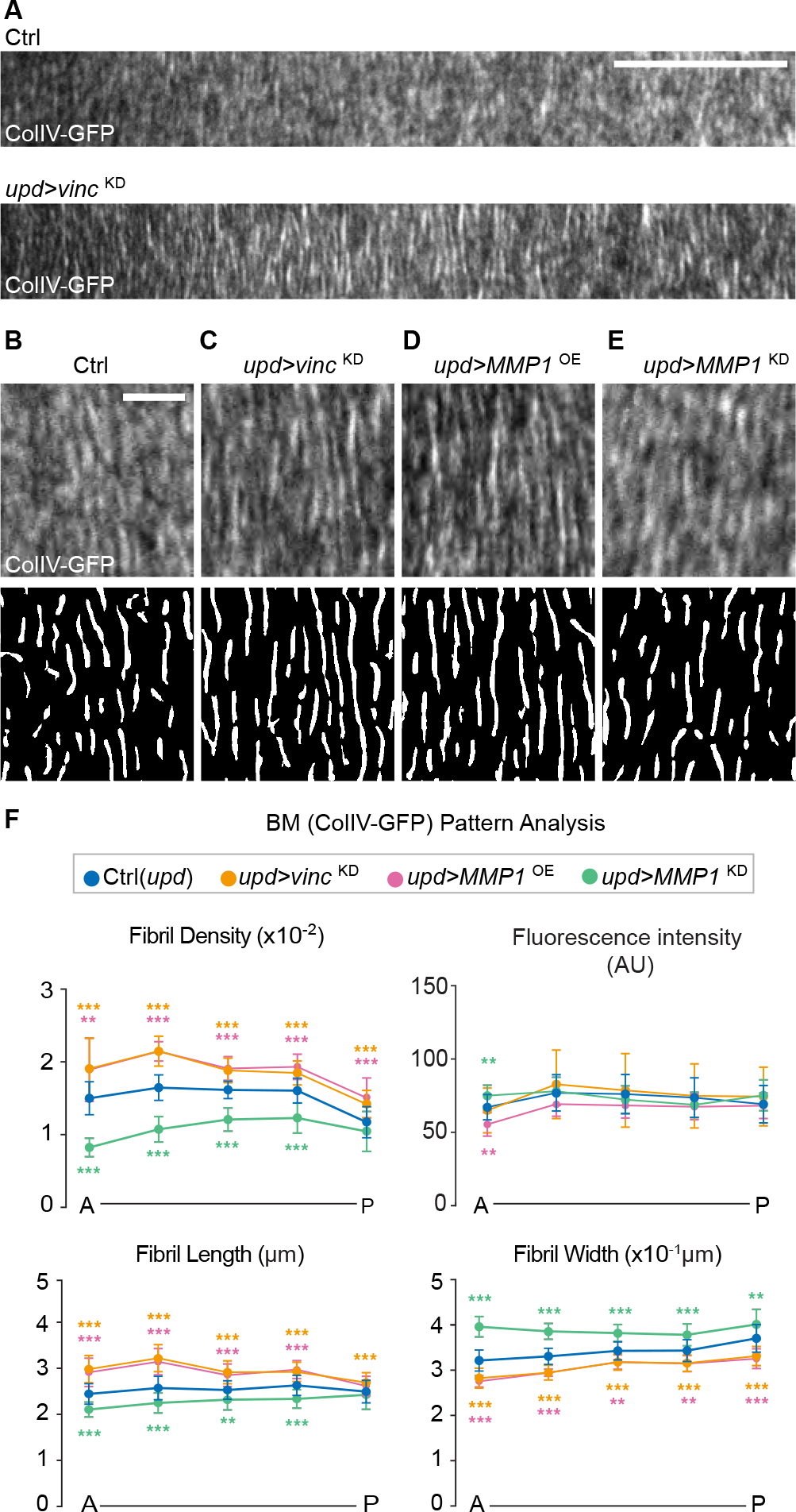
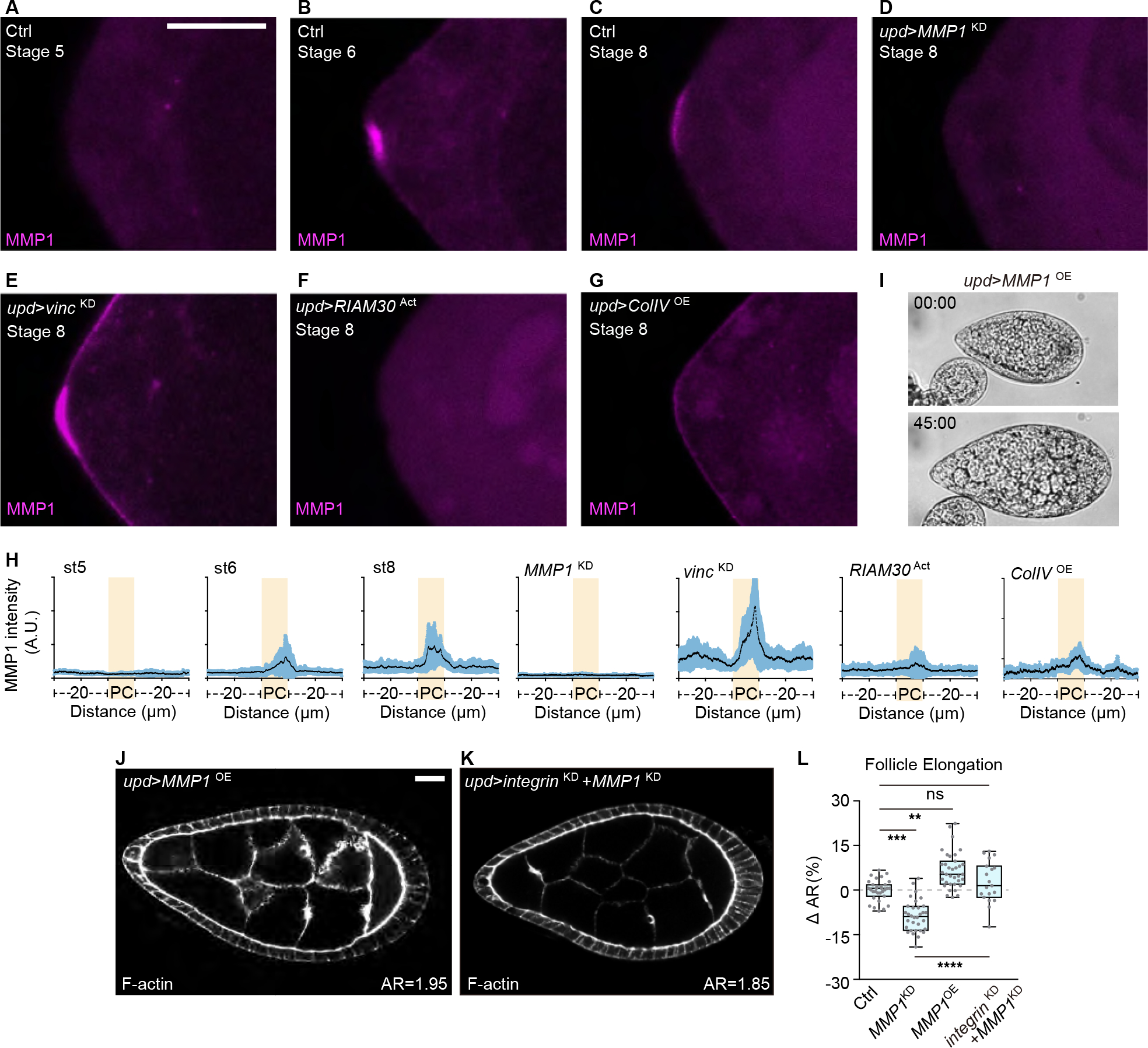
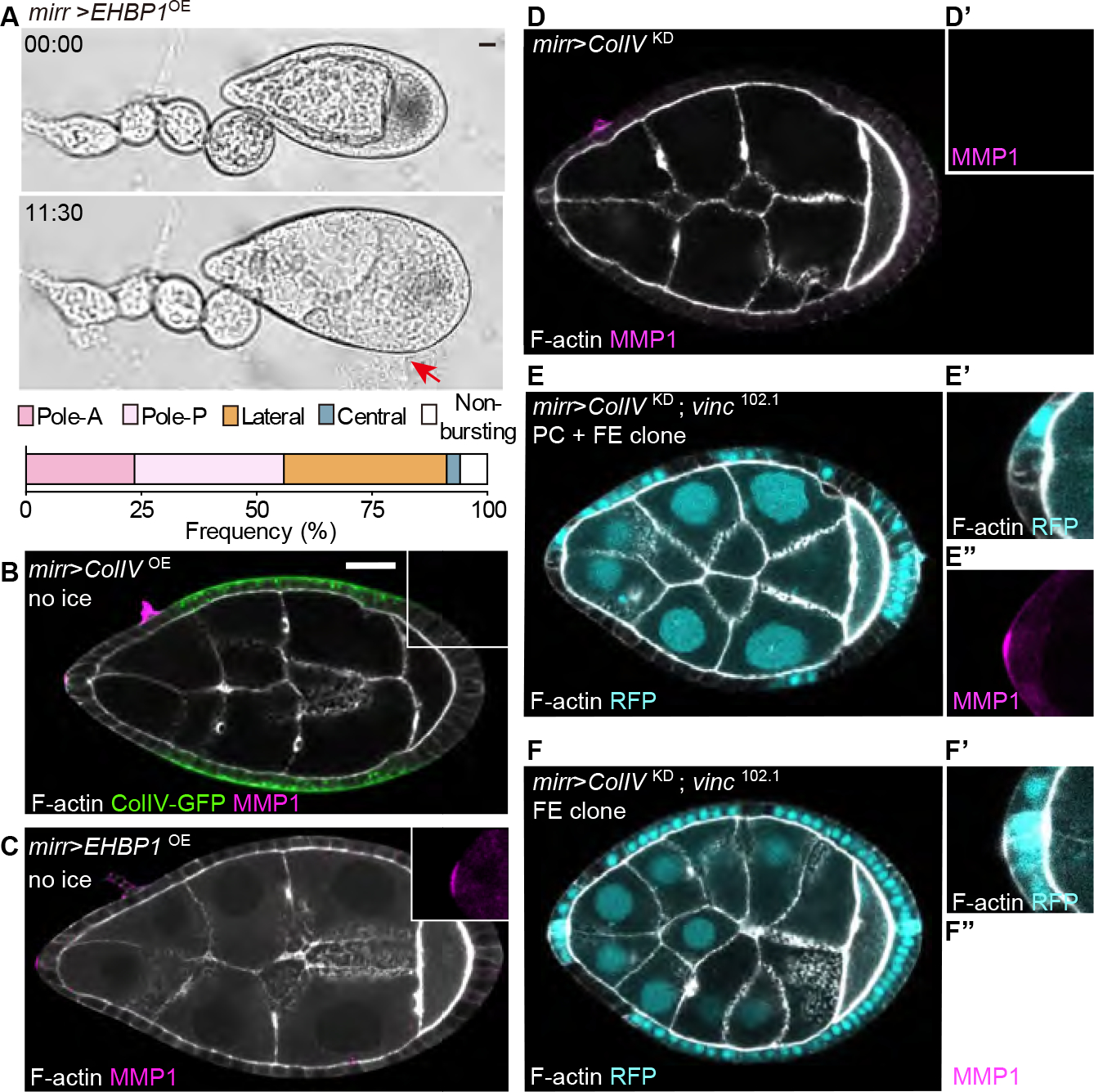
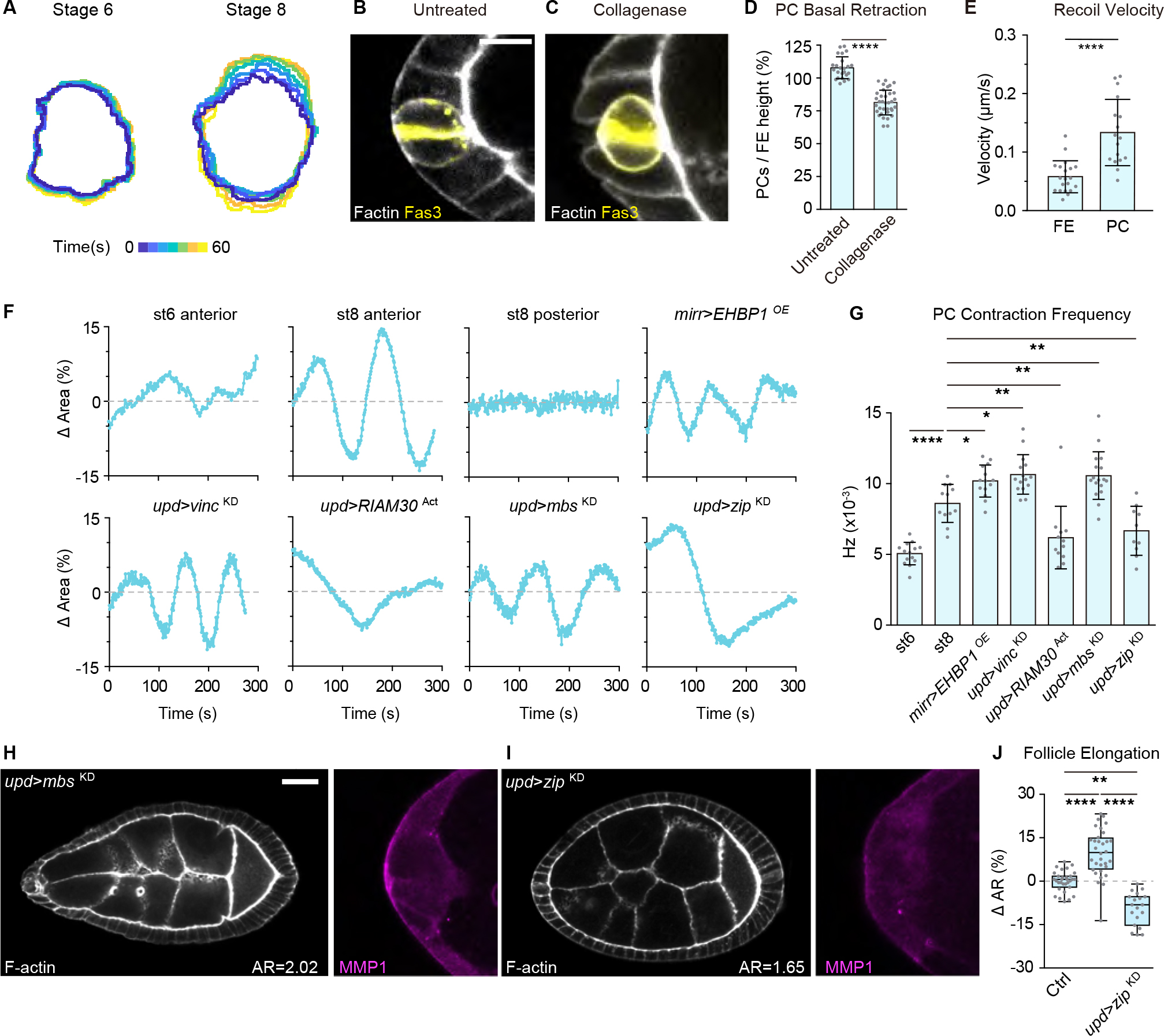
Shaping of developing organs requires dynamic regulation of force and resistance to achieve precise outcomes, but how organs monitor tissue mechanical properties is poorly understood. We show that in developing Drosophila follicles (egg chambers), a single pair of cells performs such monitoring to drive organ shaping. These anterior polar cells secrete a matrix metalloproteinase (MMP) that specifies the appropriate degree of tissue elongation, rather than hyper- or hypo-elongated organs. MMP production is negatively regulated by basement membrane (BM) mechanical properties, which are sensed through focal adhesion signaling and autonomous contractile activity; MMP then reciprocally regulates BM remodeling, particularly at the anterior region. Changing BM properties at remote locations alone is sufficient to induce a remodeling response in polar cells. We propose that this small group of cells senses both local and distant stiffness cues to produce factors that pattern the organ's BM mechanics, ensuring proper tissue shape and reproductive success.
Keywords: Drosophila; MMP; basement membrane; egg chamber; follicle; morphogenesis; tissue mechanics.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Variations in basement membrane mechanics are linked to epithelial morphogenesis.Development. 2017 Dec 1;144(23):4350-4362. doi: 10.1242/dev.152652. Epub 2017 Oct 16. Development. 2017. PMID: 29038305
-
Regulation of the collagen IV network by the basement membrane protein perlecan is crucial for squamous epithelial cell morphogenesis and organ architecture.Matrix Biol. 2022 Dec;114:35-66. doi: 10.1016/j.matbio.2022.10.004. Epub 2022 Nov 5. Matrix Biol. 2022. PMID: 36343860
-
Extracellular matrix stiffness cues junctional remodeling for 3D tissue elongation.Nat Commun. 2019 Jul 26;10(1):3339. doi: 10.1038/s41467-019-10874-x. Nat Commun. 2019. PMID: 31350387 Free PMC article.
-
Basement membrane dynamics and mechanics in tissue morphogenesis.Biol Open. 2023 Aug 15;12(8):bio059980. doi: 10.1242/bio.059980. Epub 2023 Aug 2. Biol Open. 2023. PMID: 37531197 Free PMC article. Review.
-
ECM-modulated cellular dynamics as a driving force for tissue morphogenesis.Curr Opin Genet Dev. 2013 Aug;23(4):408-14. doi: 10.1016/j.gde.2013.05.005. Epub 2013 Jul 9. Curr Opin Genet Dev. 2013. PMID: 23849799 Free PMC article. Review.
Cited by
-
Bellymount-pulsed tracking: a novel approach for real-time in vivo imaging of Drosophila abdominal tissues.G3 (Bethesda). 2025 Jan 8;15(1):jkae271. doi: 10.1093/g3journal/jkae271. G3 (Bethesda). 2025. PMID: 39556480 Free PMC article.
-
The life cycle of type IV collagen.Matrix Biol. 2025 Aug;139:14-28. doi: 10.1016/j.matbio.2025.04.004. Epub 2025 Apr 28. Matrix Biol. 2025. PMID: 40306374 Review.
-
A complex relationship between the architecture of the basement membrane, its mechanical properties, and its ability to shape the Drosophila egg.Matrix Biol. 2025 Sep;140:16-26. doi: 10.1016/j.matbio.2025.06.001. Epub 2025 Jun 13. Matrix Biol. 2025. PMID: 40518025
-
Basement membrane patterning by spatial deployment of a secretion-regulating protease.Proc Natl Acad Sci U S A. 2025 May 20;122(20):e2412161122. doi: 10.1073/pnas.2412161122. Epub 2025 May 13. Proc Natl Acad Sci U S A. 2025. PMID: 40359035
-
A mechanism-based theory of cellular and tissue plasticity.Proc Natl Acad Sci U S A. 2023 Oct 31;120(44):e2305375120. doi: 10.1073/pnas.2305375120. Epub 2023 Oct 23. Proc Natl Acad Sci U S A. 2023. PMID: 37871208 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
