

Oxytocin receptor is not required for social attachment in prairie voles
- PMID: 36708707
- PMCID: PMC10150797
- DOI: 10.1016/j.neuron.2022.12.011
Oxytocin receptor is not required for social attachment in prairie voles
Abstract
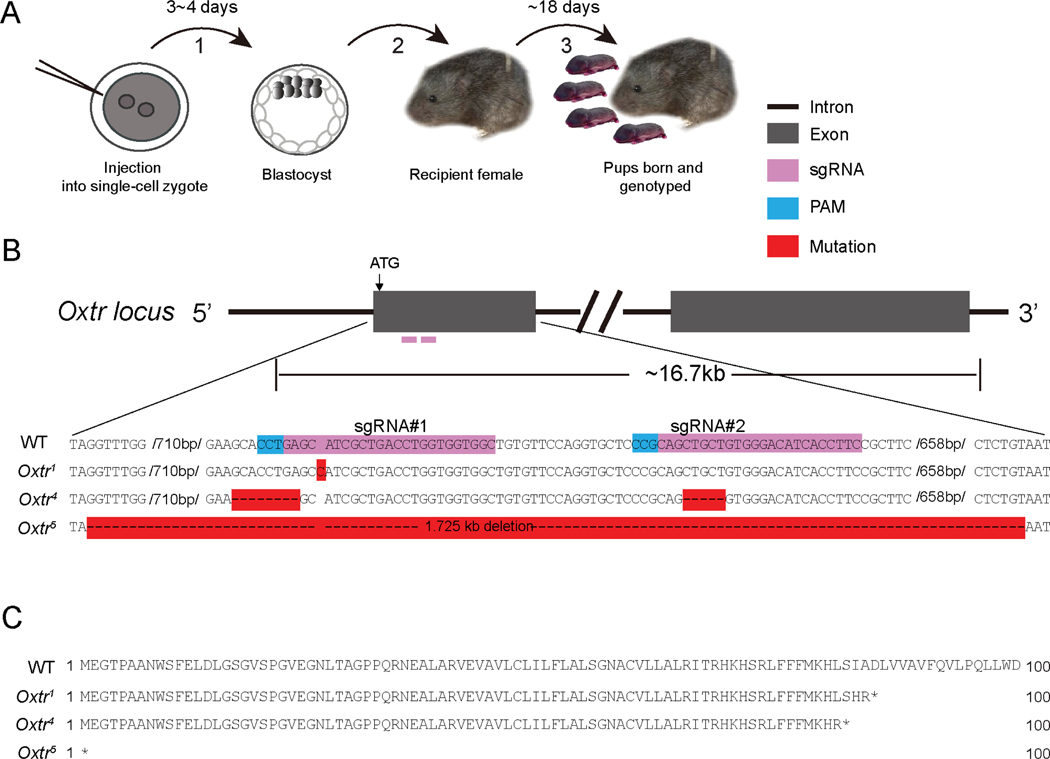
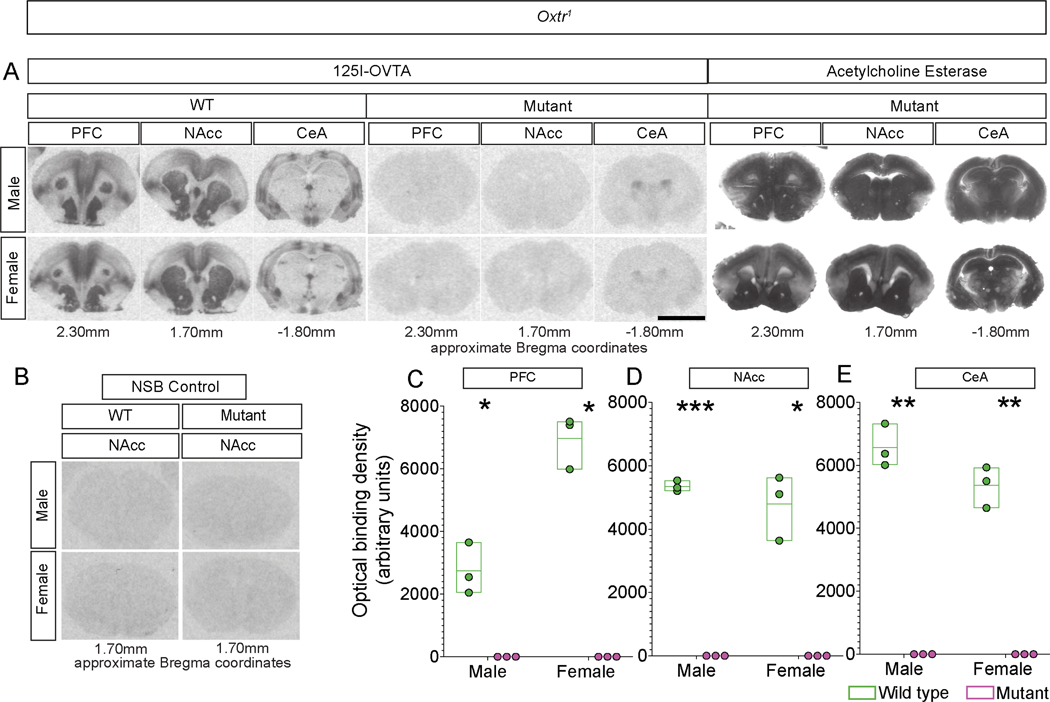
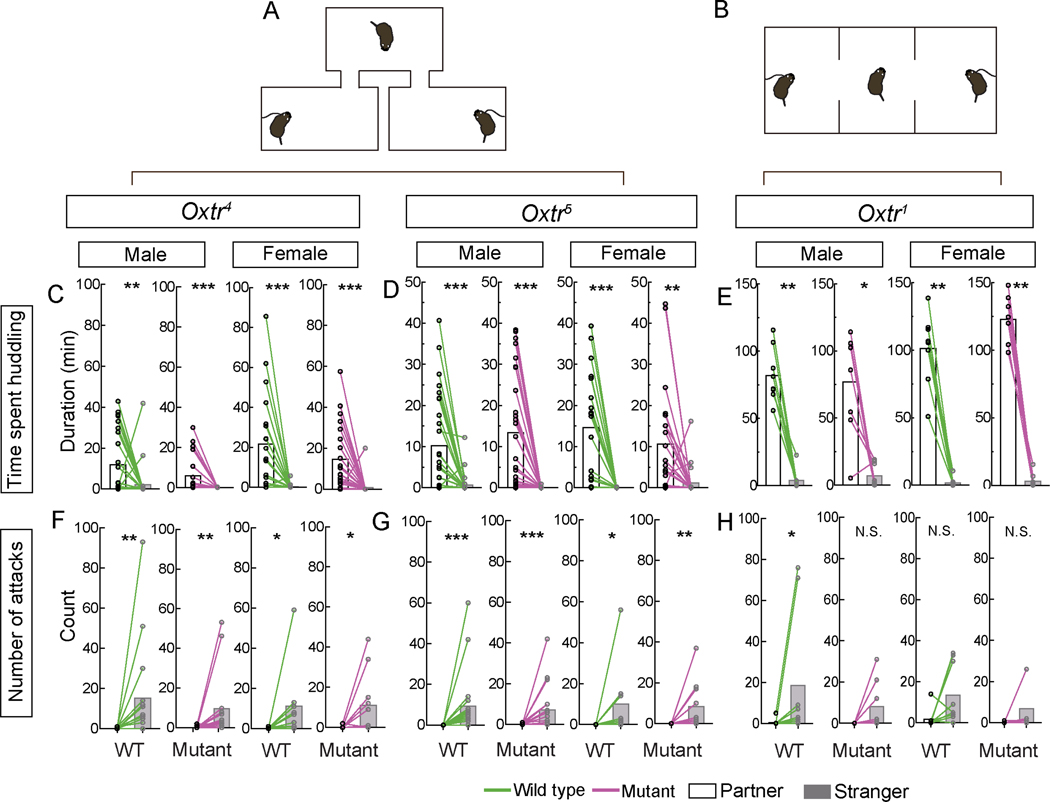
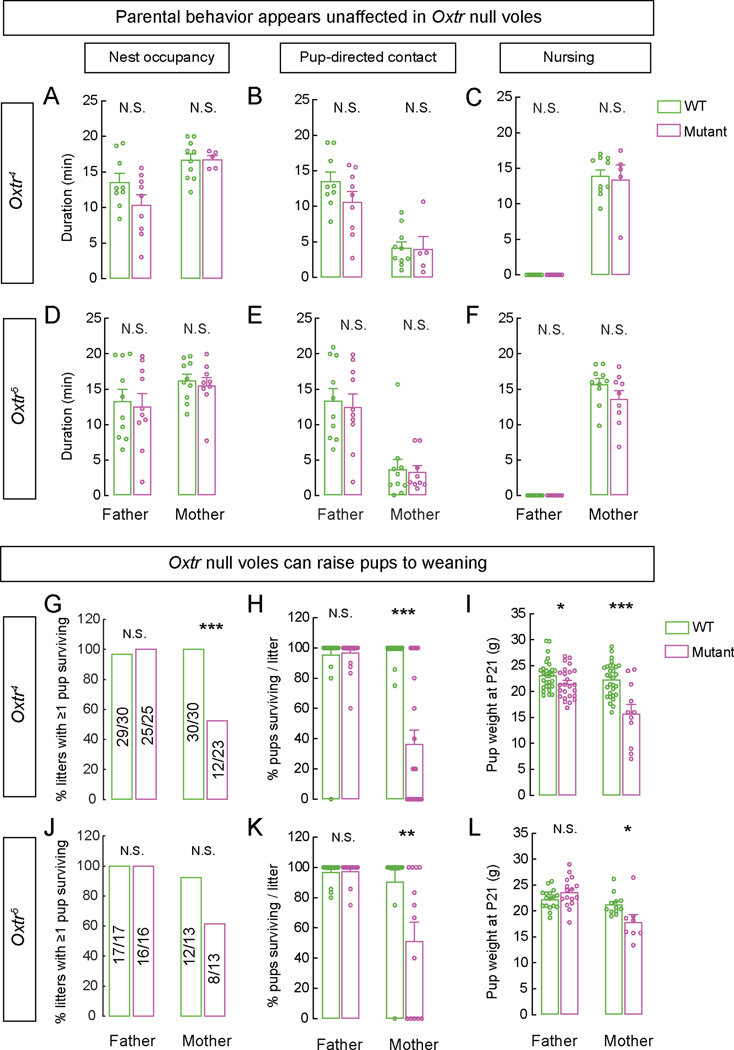
Prairie voles are among a small group of mammals that display long-term social attachment between mating partners. Many pharmacological studies show that signaling via the oxytocin receptor (Oxtr) is critical for the display of social monogamy in these animals. We used CRISPR mutagenesis to generate three different Oxtr-null mutant prairie vole lines. Oxtr mutants displayed social attachment such that males and females showed a behavioral preference for their mating partners over a stranger of the opposite sex, even when assayed using different experimental setups. Mothers lacking Oxtr delivered viable pups, and parents displayed care for their young and raised them to the weanling stage. Together, our studies unexpectedly reveal that social attachment, parturition, and parental behavior can occur in the absence of Oxtr signaling in prairie voles.
Keywords: CRISPR; monogamy; nursing; oxytocin receptor; pair-bonding; parental behavior; partner preference; prairie vole; social attachment.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Dr. Nirao Shah is a member of the advisory board for Neuron.
Figures
Comment in
-
A CRISPR perspective of the oxytocin receptor in prairie voles.Neuron. 2023 Mar 15;111(6):755-756. doi: 10.1016/j.neuron.2023.02.011. Neuron. 2023. PMID: 36924758
