

Seasonal cues act through the circadian clock and pigment-dispersing factor to control EYES ABSENT and downstream physiological changes
- PMID: 36708710
- PMCID: PMC9992282
- DOI: 10.1016/j.cub.2023.01.006
Seasonal cues act through the circadian clock and pigment-dispersing factor to control EYES ABSENT and downstream physiological changes
Erratum in
-
Seasonal cues act through the circadian clock and pigment-dispersing factor to control EYES ABSENT and downstream physiological changes.Curr Biol. 2023 Mar 13;33(5):1006-1008. doi: 10.1016/j.cub.2023.02.028. Curr Biol. 2023. PMID: 36917930 Free PMC article. No abstract available.
Abstract
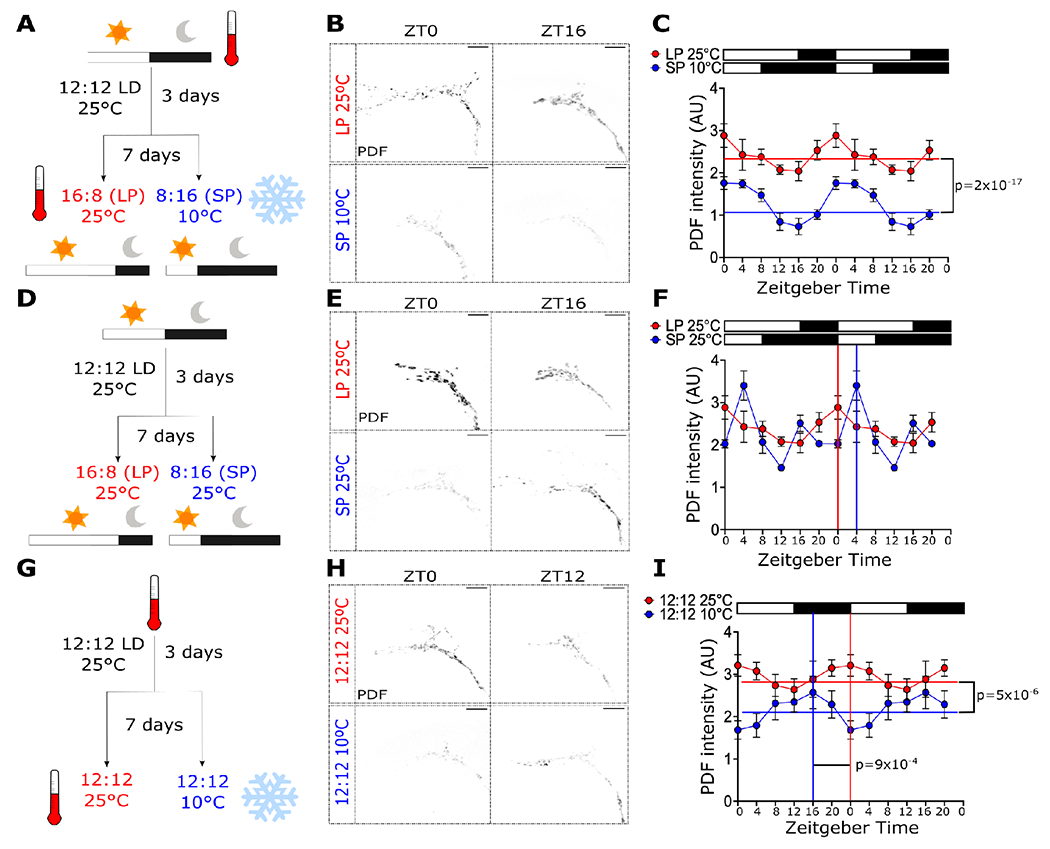
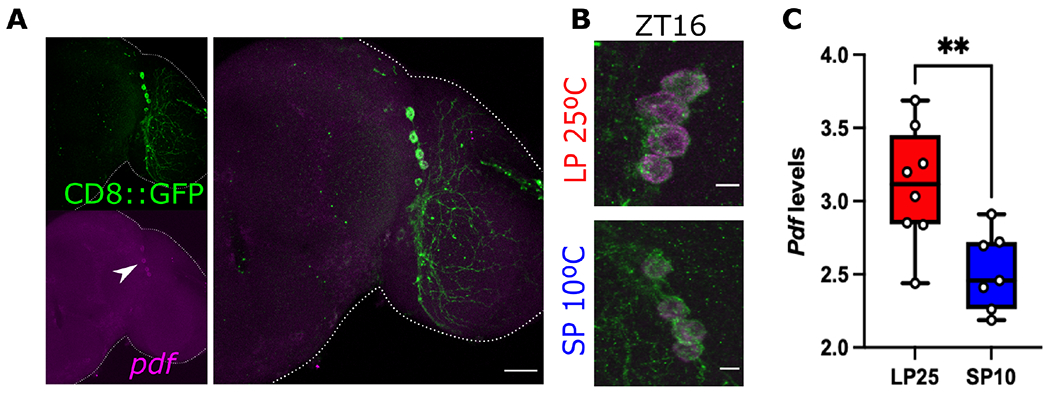
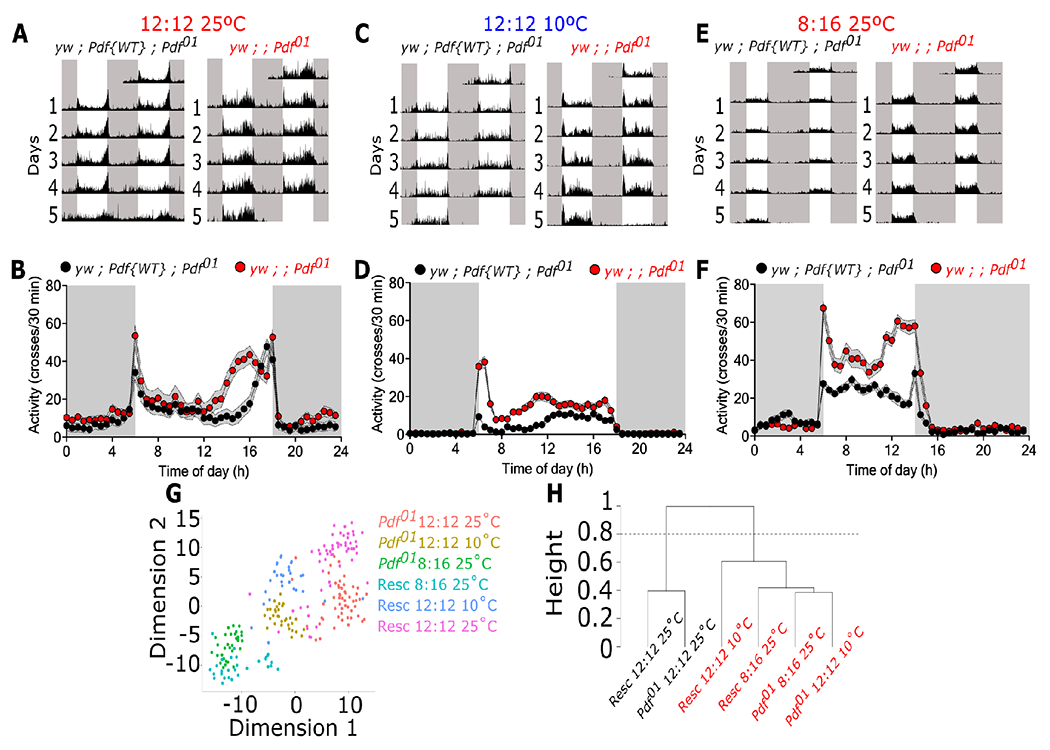
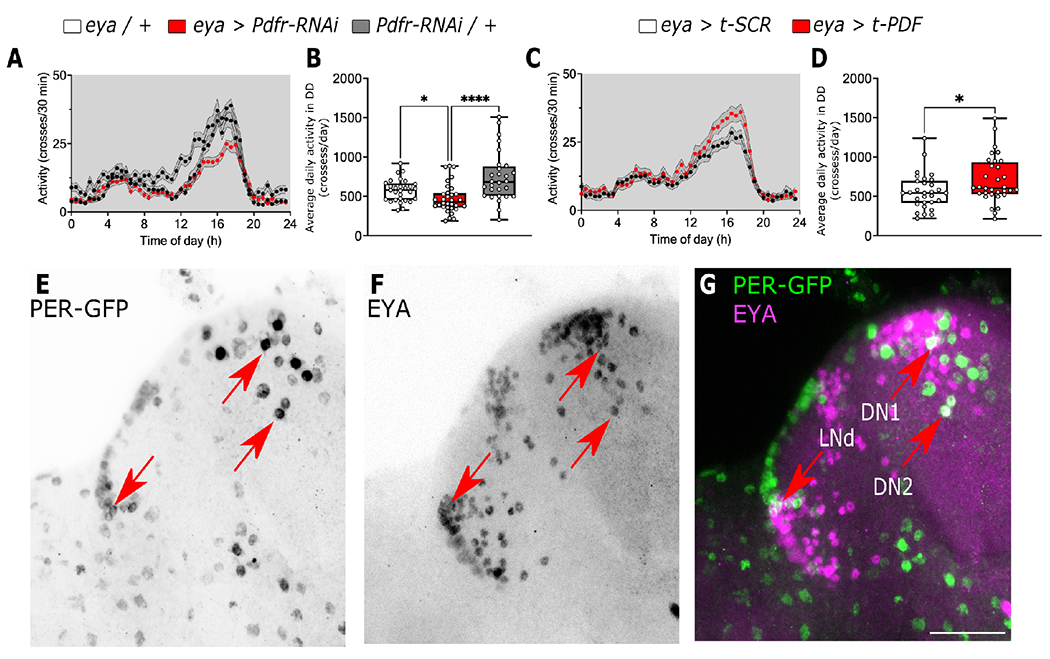
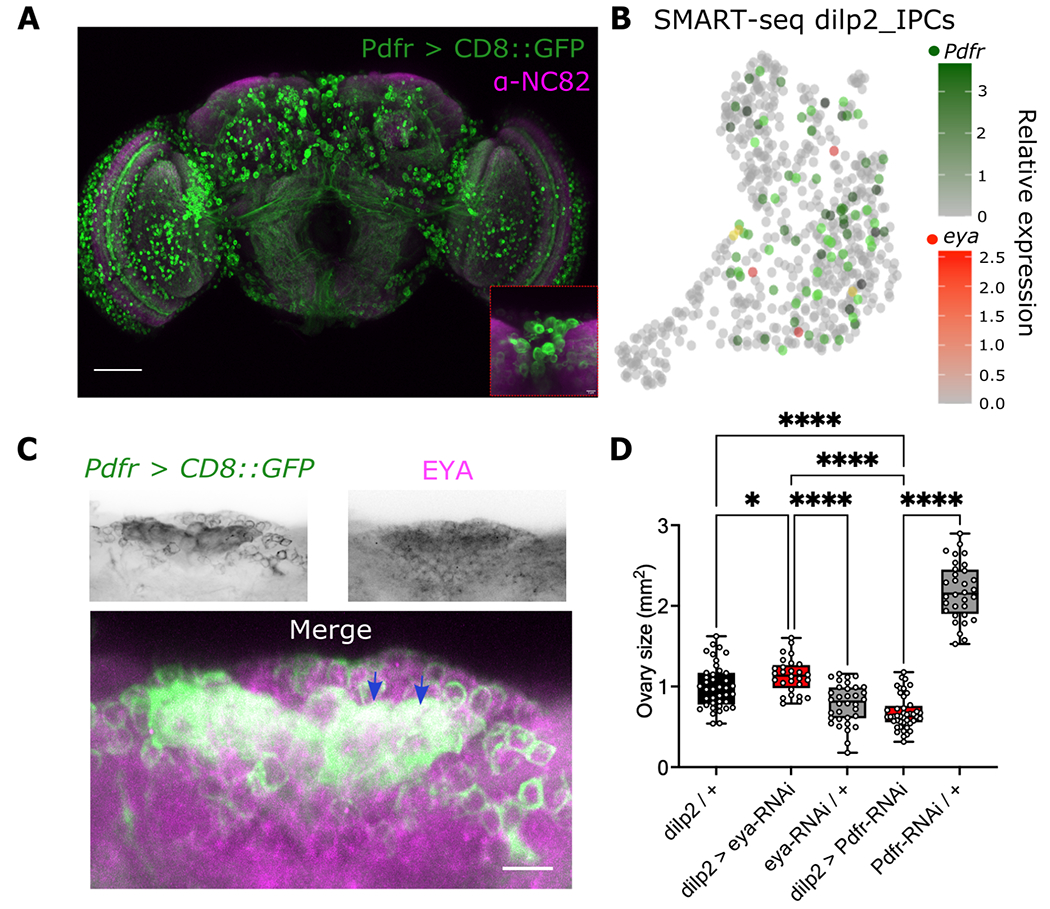
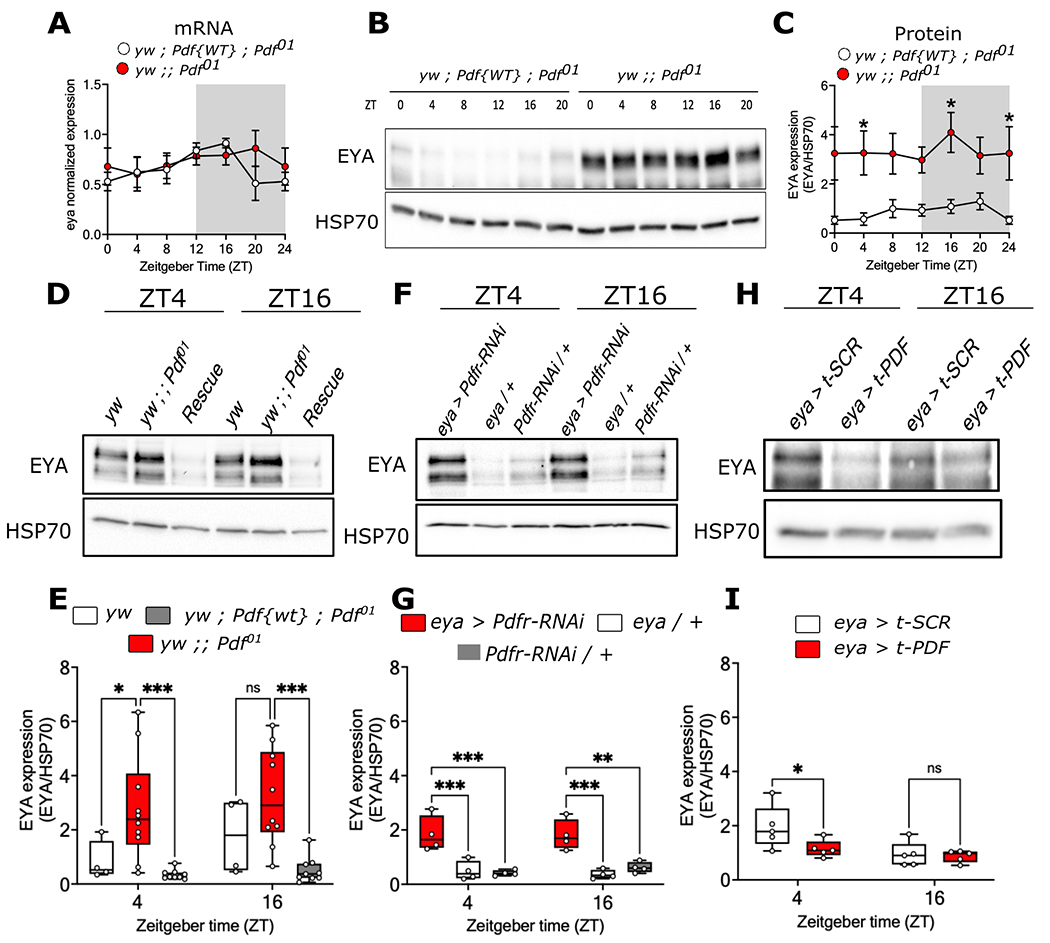
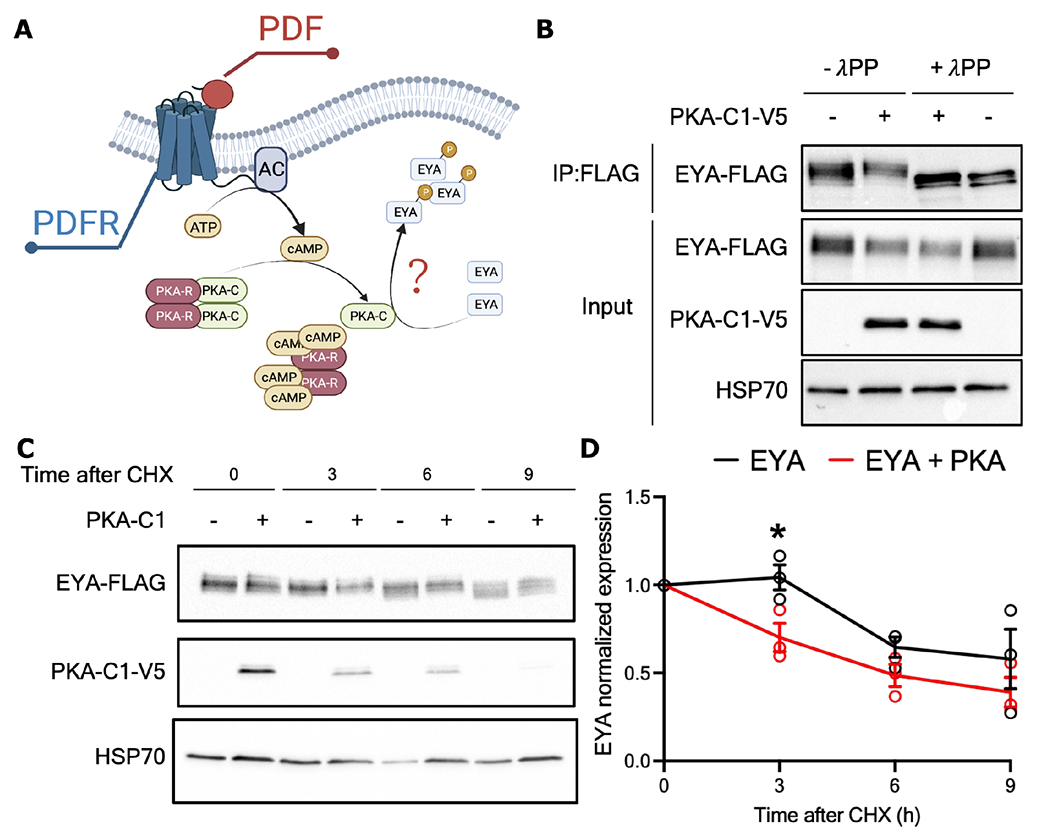
Organisms adapt to seasonal changes in photoperiod and temperature to survive; however, the mechanisms by which these signals are integrated in the brain to alter seasonal biology are poorly understood. We previously reported that EYES ABSENT (EYA) shows higher levels in cold temperature or short photoperiod and promotes winter physiology in Drosophila. Nevertheless, how EYA senses seasonal cues is unclear. Pigment-dispersing factor (PDF) is a neuropeptide important for regulating circadian output rhythms. Interestingly, PDF has also been shown to regulate seasonality, suggesting that it may mediate the function of the circadian clock in modulating seasonal physiology. In this study, we investigated the role of EYA in mediating the function of PDF on seasonal biology. We observed that PDF abundance is lower on cold and short days as compared with warm and long days, contrary to what was previously observed for EYA. We observed that manipulating PDF signaling in eya+ fly brain neurons, where EYA and PDF receptor are co-expressed, modulates seasonal adaptations in daily activity rhythm and ovary development via EYA-dependent and EYA-independent mechanisms. At the molecular level, altering PDF signaling impacted EYA protein abundance. Specifically, we showed that protein kinase A (PKA), an effector of PDF signaling, phosphorylates EYA promoting its degradation, thus explaining the opposite responses of PDF and EYA abundance to changes in seasonal cues. In summary, our results support a model in which PDF signaling negatively modulates EYA levels to regulate seasonal physiology, linking the circadian clock to the modulation of seasonal adaptations.
Keywords: circadian clock; daily activity rhythm; insect reproductive dormancy; neuropeptide; photoperiod; post-translational modification; seasonal biology; temperature.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Biological timing: Linking the circadian clock to the season.Curr Biol. 2023 Feb 27;33(4):R141-R143. doi: 10.1016/j.cub.2023.01.026. Curr Biol. 2023. PMID: 36854270
Similar articles
-
EYES ABSENT and TIMELESS integrate photoperiodic and temperature cues to regulate seasonal physiology in Drosophila.Proc Natl Acad Sci U S A. 2020 Jun 30;117(26):15293-15304. doi: 10.1073/pnas.2004262117. Epub 2020 Jun 15. Proc Natl Acad Sci U S A. 2020. PMID: 32541062 Free PMC article.
-
Integration of photoperiodic and temperature cues by the circadian clock to regulate insect seasonal adaptations.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024 Jul;210(4):585-599. doi: 10.1007/s00359-023-01667-1. Epub 2023 Aug 16. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024. PMID: 37584703 Free PMC article. Review.
-
Regulation of Olfactory Associative Memory by the Circadian Clock Output Signal Pigment-Dispersing Factor (PDF).J Neurosci. 2020 Nov 18;40(47):9066-9077. doi: 10.1523/JNEUROSCI.0782-20.2020. Epub 2020 Oct 26. J Neurosci. 2020. PMID: 33106351 Free PMC article.
-
Phase coupling of a circadian neuropeptide with rest/activity rhythms detected using a membrane-tethered spider toxin.PLoS Biol. 2008 Nov 4;6(11):e273. doi: 10.1371/journal.pbio.0060273. PLoS Biol. 2008. PMID: 18986214 Free PMC article.
-
Downloading central clock information in Drosophila.Mol Neurobiol. 2002 Oct-Dec;26(2-3):217-33. doi: 10.1385/MN:26:2-3:217. Mol Neurobiol. 2002. PMID: 12428757 Review.
Cited by
-
Correlation between circadian and photoperiodic latitudinal clines in Drosophila littoralis.Open Biol. 2025 Mar;15(3):240403. doi: 10.1098/rsob.240403. Epub 2025 Mar 5. Open Biol. 2025. PMID: 40037533 Free PMC article.
-
Synaptic Targets of Circadian Clock Neurons Influence Core Clock Parameters.bioRxiv [Preprint]. 2025 Feb 6:2025.01.30.635801. doi: 10.1101/2025.01.30.635801. bioRxiv. 2025. PMID: 39975067 Free PMC article. Preprint.
-
One seasonal clock fits all?J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024 Jul;210(4):641-647. doi: 10.1007/s00359-023-01680-4. Epub 2023 Nov 10. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024. PMID: 37947808 Free PMC article. Review.
-
The circadian and photoperiodic clock of the pea aphid.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024 Jul;210(4):627-639. doi: 10.1007/s00359-023-01660-8. Epub 2023 Jul 24. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024. PMID: 37482577 Free PMC article. Review.
-
Pigment-dispersing factor is present in circadian clock neurons of pea aphids and may mediate photoperiodic signalling to insulin-producing cells.Open Biol. 2023 Jun;13(6):230090. doi: 10.1098/rsob.230090. Epub 2023 Jun 28. Open Biol. 2023. PMID: 37369351 Free PMC article.
References
-
- Stevenson TJ, Prendergast BJ, and Nelson RJ (2017). Mammalian Seasonal Rhythms: Behavior and Neuroendocrine Substrates. In Hormones, Brain and Behavior: Third edition (Academic Press; ), pp. 371–398. 10.1016/B978-0-12-803592-4.00013-4. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
