

This is a preprint.
RNA Polymerase II, the BAF remodeler and transcription factors synergize to evict nucleosomes
- PMID: 36711459
- PMCID: PMC9882304
- DOI: 10.1101/2023.01.22.525083
RNA Polymerase II, the BAF remodeler and transcription factors synergize to evict nucleosomes
Update in
-
The BAF chromatin remodeler synergizes with RNA polymerase II and transcription factors to evict nucleosomes.Nat Genet. 2024 Jan;56(1):100-111. doi: 10.1038/s41588-023-01603-8. Epub 2023 Dec 4. Nat Genet. 2024. PMID: 38049663 Free PMC article.
Abstract
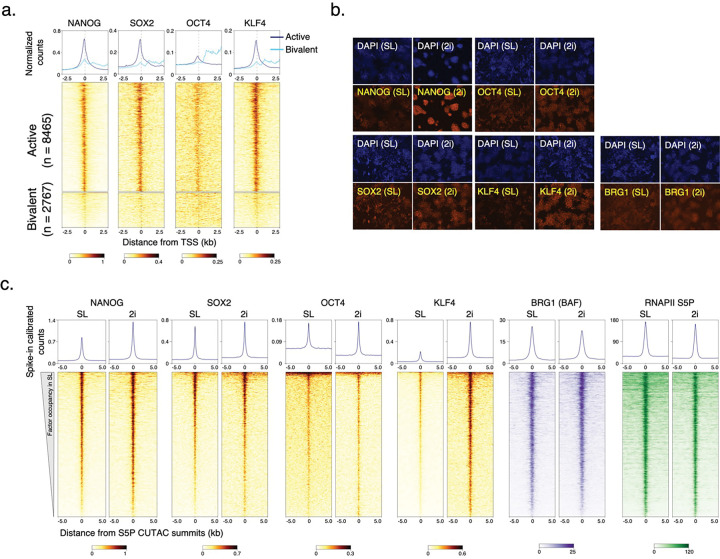
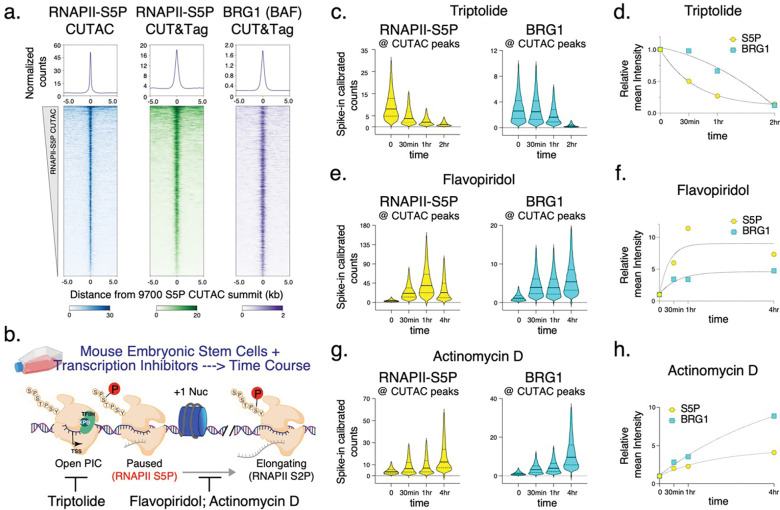
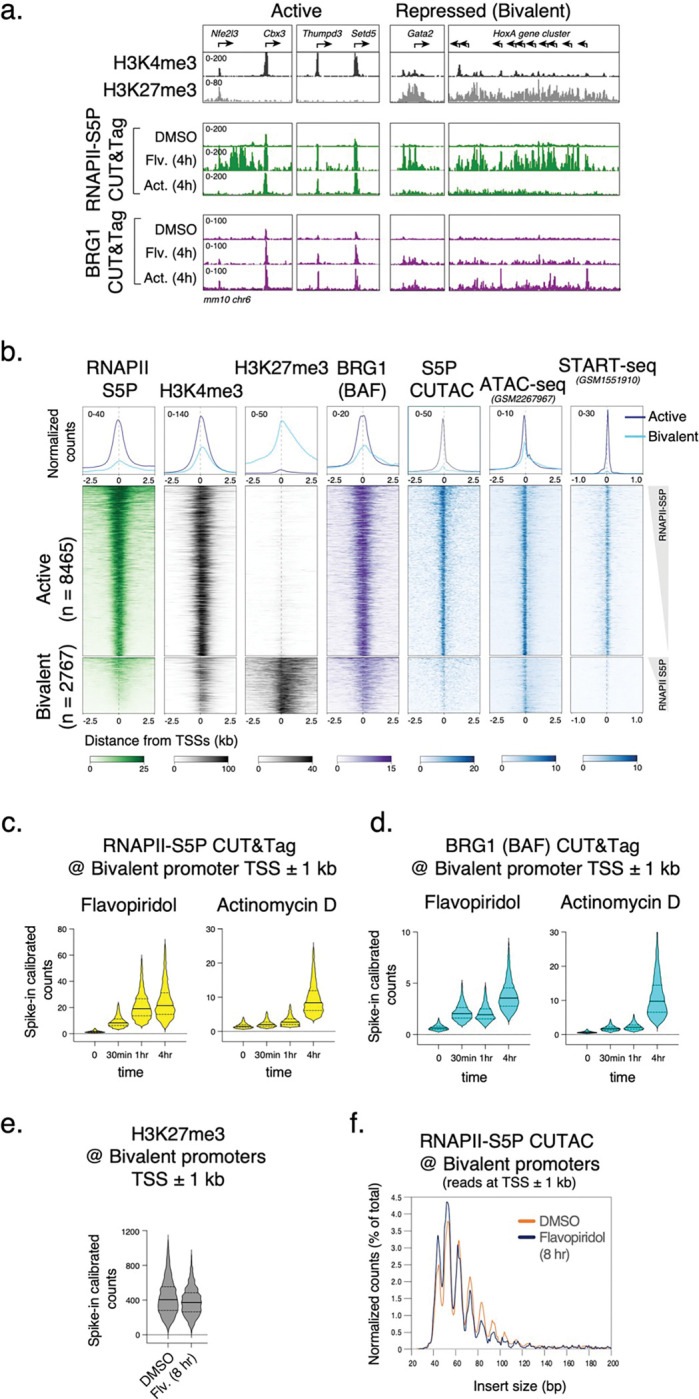
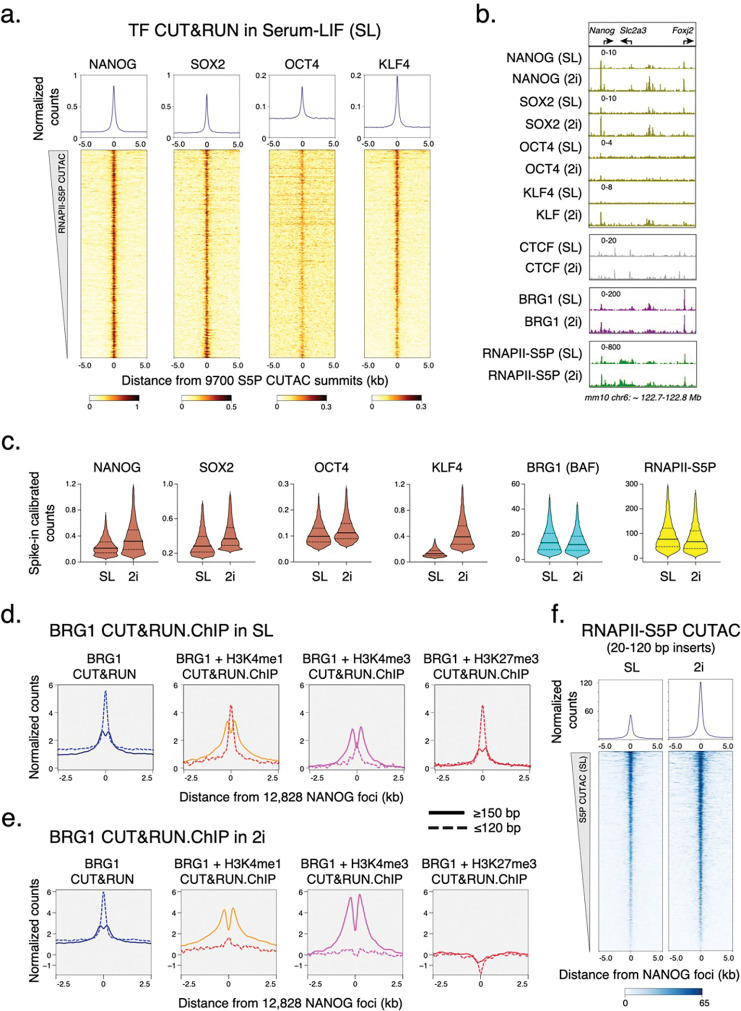
Chromatin accessibility is a hallmark of active transcription and requires ATP-dependent nucleosome remodeling by Brahma-Associated Factor (BAF). However, the mechanistic link between transcription, nucleosome remodeling, and chromatin accessibility is unclear. Here, we used a chemical-genetic approach to dissect the interplay between RNA Polymerase II (RNAPII), BAF, and DNA-sequence-specific transcription factors (TFs) in mouse embryonic stem cells. By time-resolved chromatin profiling with acute transcription block at distinct stages, we show that RNAPII promoter-proximal pausing stabilizes BAF chromatin occupancy and enhances nucleosome eviction by BAF. We find that RNAPII and BAF probe both transcriptionally active and Polycomb-repressed genomic regions and provide evidence that TFs capture transient site exposure due to nucleosome unwrapping by BAF to confer locus specificity for persistent chromatin remodeling. Our study reveals the mechanistic basis of cell-type-specific chromatin accessibility. We propose a new paradigm for how functional synergy between dynamically acting chromatin factors regulates nucleosome organization.
Conflict of interest statement
Declaration of interest The authors declare no competing interests.
Figures
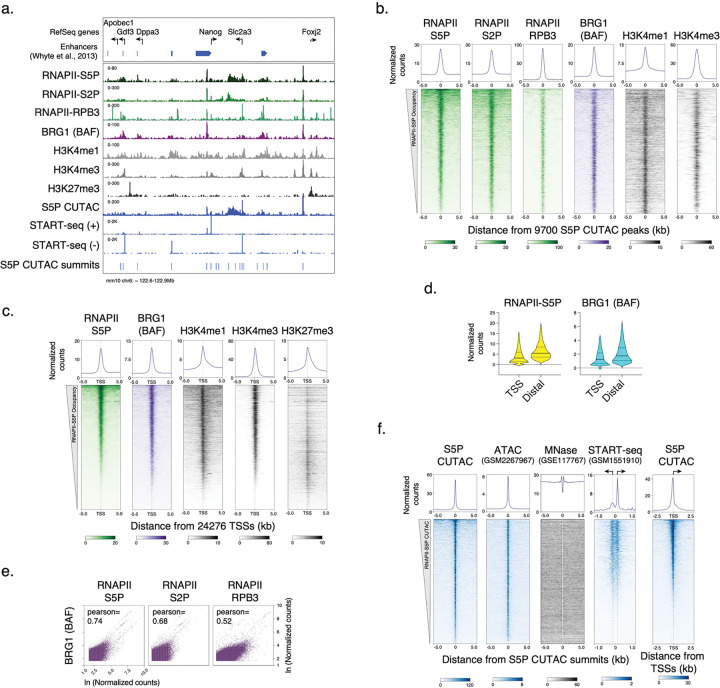
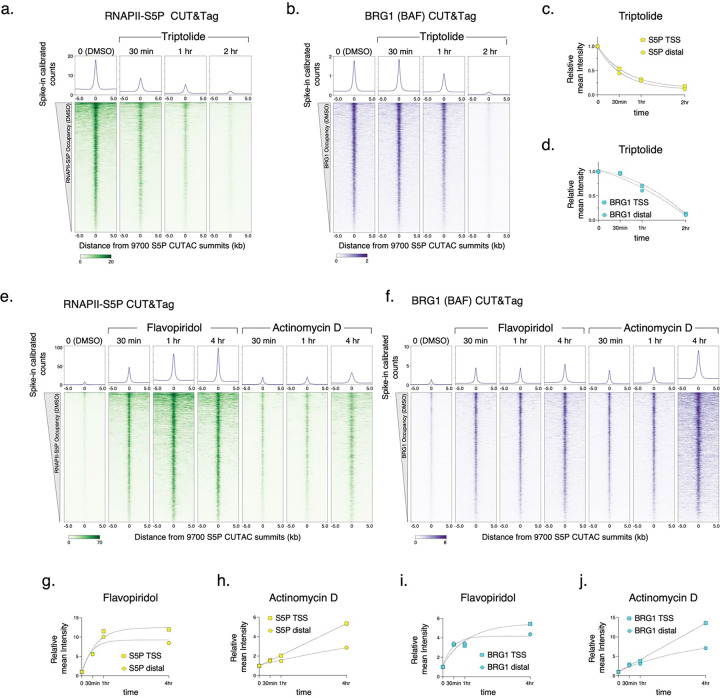
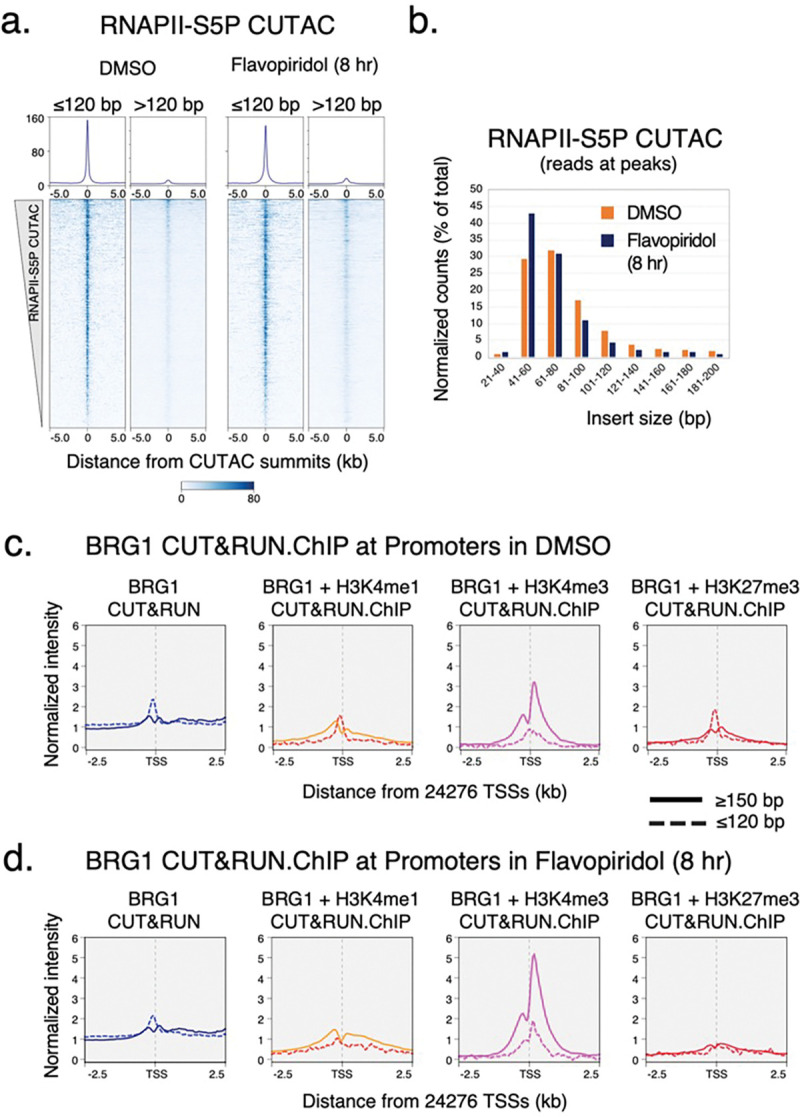
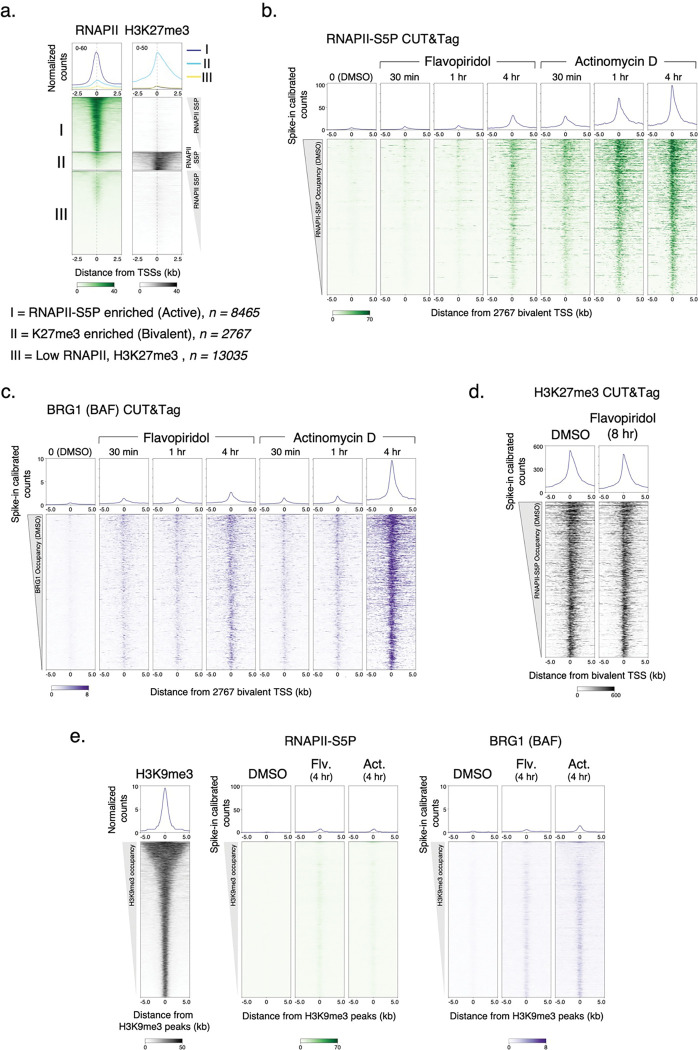


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources