

This is a preprint.
Maturation of glutamatergic transmission onto dorsal raphe serotonergic neurons
- PMID: 36711665
- PMCID: PMC9882295
- DOI: 10.1101/2023.01.19.524776
Maturation of glutamatergic transmission onto dorsal raphe serotonergic neurons
Update in
-
Maturation of glutamatergic transmission onto dorsal raphe serotonergic neurons.J Neurophysiol. 2024 Apr 1;131(4):626-637. doi: 10.1152/jn.00037.2023. Epub 2024 Feb 21. J Neurophysiol. 2024. PMID: 38380827 Free PMC article.
Abstract
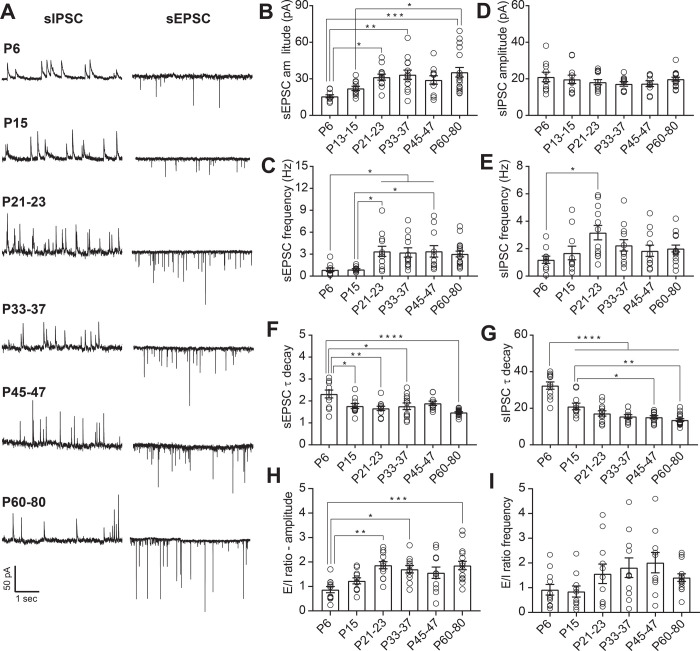
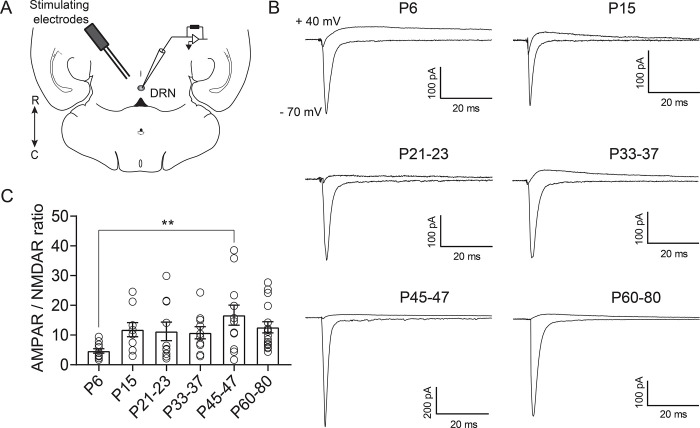
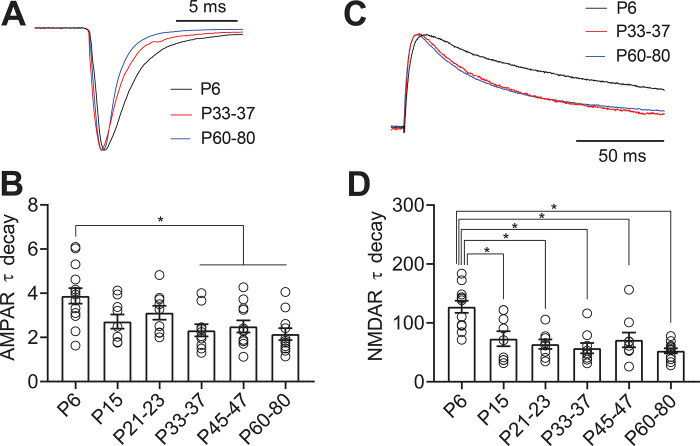
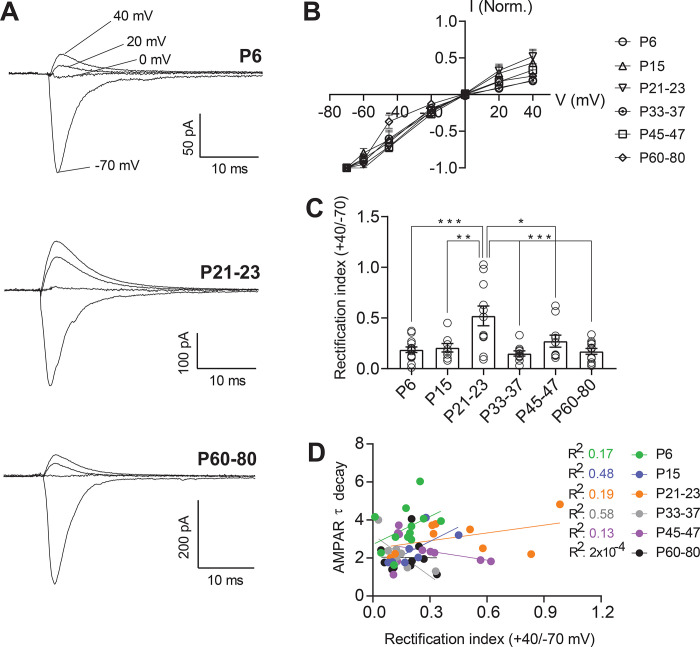
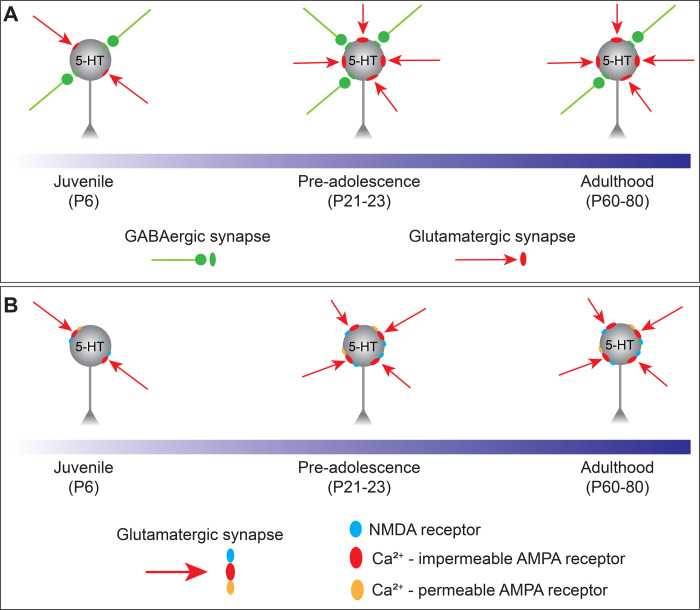
Serotonergic neurons in the dorsal raphe nucleus (DRN) play important roles early in postnatal development in the maturation and modulation of higher order emotional, sensory, and cognitive circuitry. This unique position makes these cells a substrate by which early experience can be wired into brain. In this study, we have investigated the maturation of synapses onto dorsal raphe serotonergic neurons in typically developing male and female mice using whole-cell patch-clamp recordings in ex vivo brain slices. We show that while inhibition of these neurons is relatively stable across development, glutamatergic synapses greatly increase in strength between P6 and P21-23. In contrast to forebrain regions, where the components making up glutamatergic synapses are dynamic across early life, we find that the makeup of these synapses onto DRN serotonergic neurons is largely stable after P15. DRN excitatory synapses maintain a very high ratio of AMPA to NMDA receptors and a rectifying component of the AMPA response throughout the lifespan. Overall, these findings reveal that the development of serotonergic neurons is marked by a significant refinement of glutamatergic synapses during the first 3 postnatal weeks. This suggests this time as a sensitive period of heightened plasticity for integration of information from upstream brain areas and that genetic and environmental insults during this period could lead to alterations in serotonergic output, impacting both the development of forebrain circuits and lifelong neuromodulatory actions.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials