

This is a preprint.
Post-fast refeeding enhances intestinal stem cell-mediated regeneration and tumourigenesis through mTORC1-dependent polyamine synthesis
- PMID: 36711807
- PMCID: PMC9882602
- DOI: 10.21203/rs.3.rs-2320717/v1
Post-fast refeeding enhances intestinal stem cell-mediated regeneration and tumourigenesis through mTORC1-dependent polyamine synthesis
Update in
-
Short-term post-fast refeeding enhances intestinal stemness via polyamines.Nature. 2024 Sep;633(8031):895-904. doi: 10.1038/s41586-024-07840-z. Epub 2024 Aug 21. Nature. 2024. PMID: 39169180 Free PMC article.
Abstract
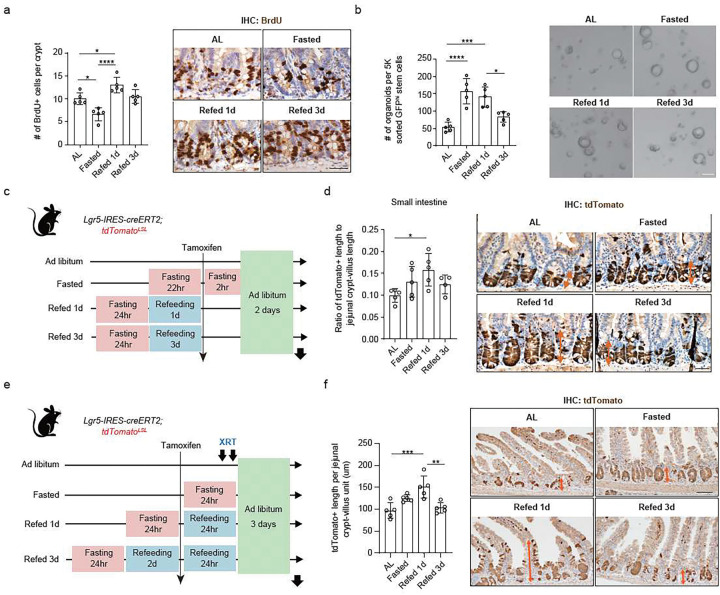
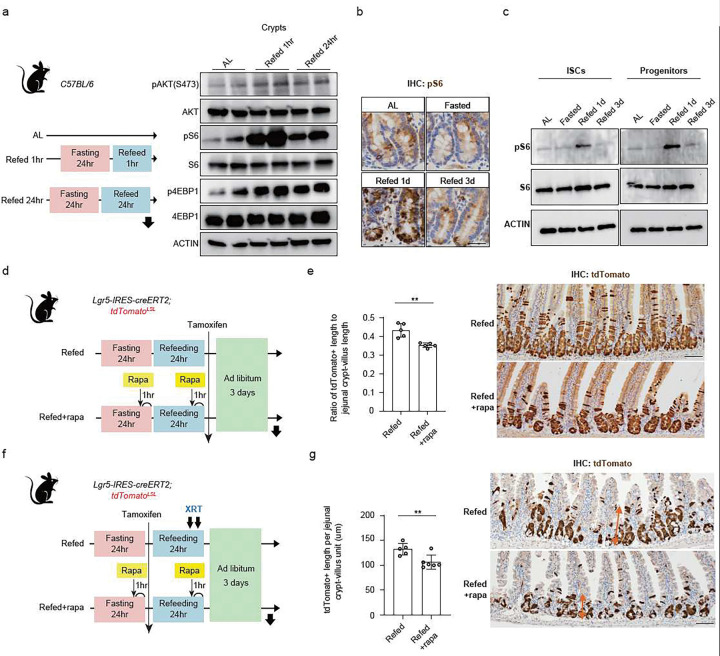
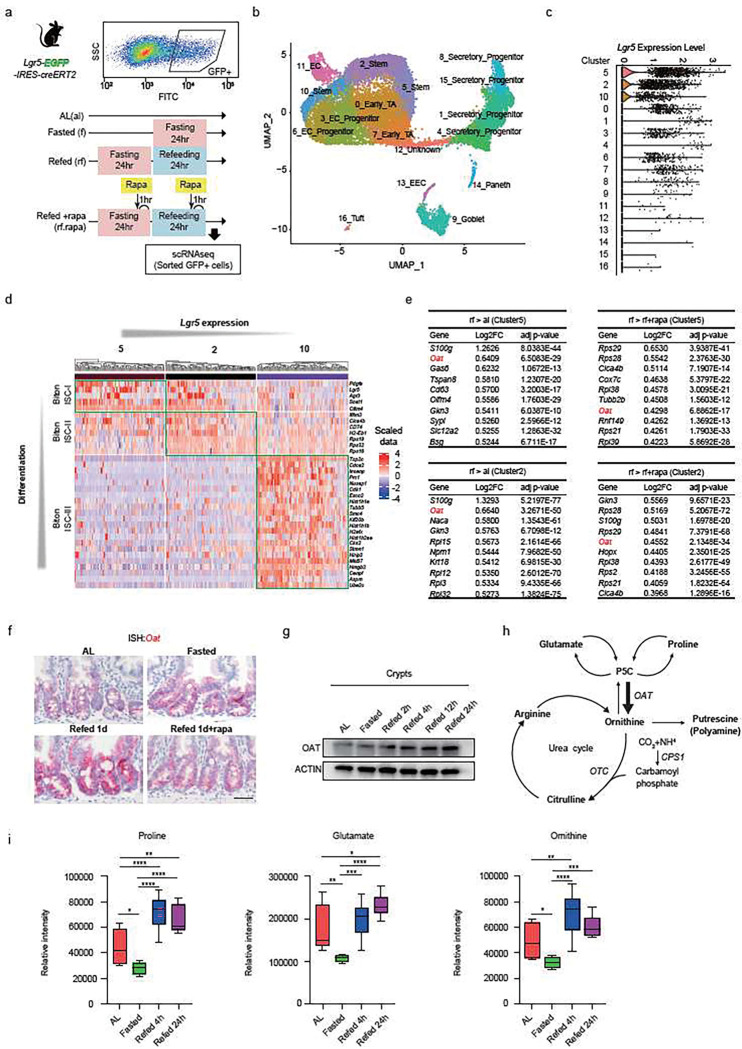
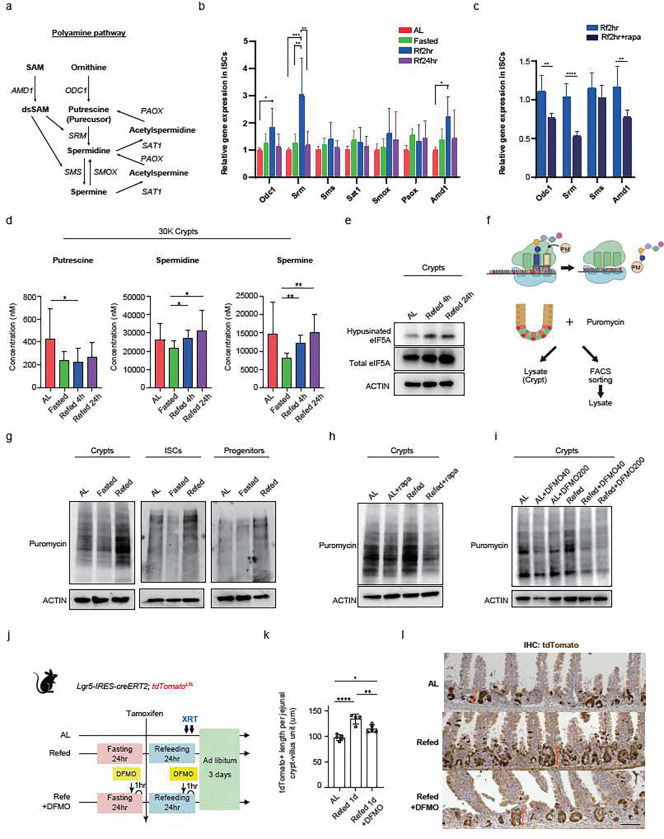
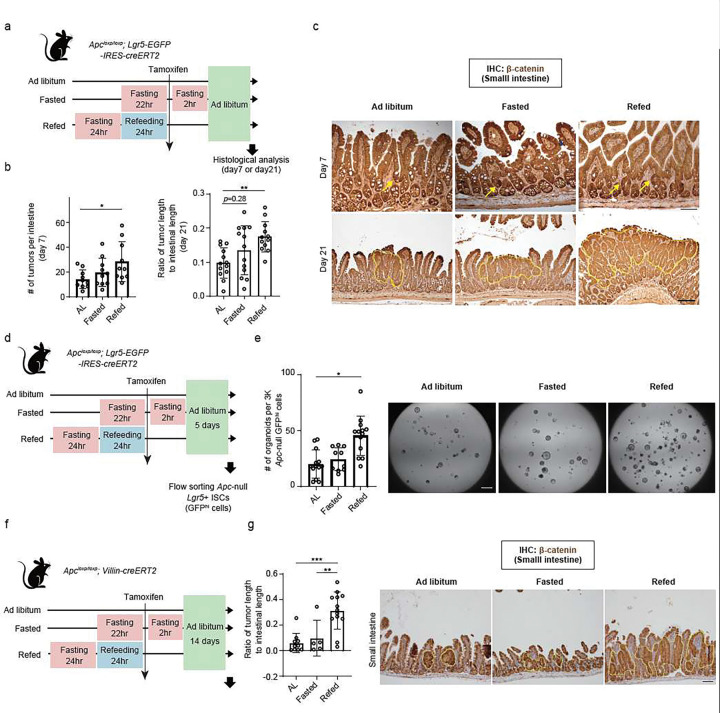
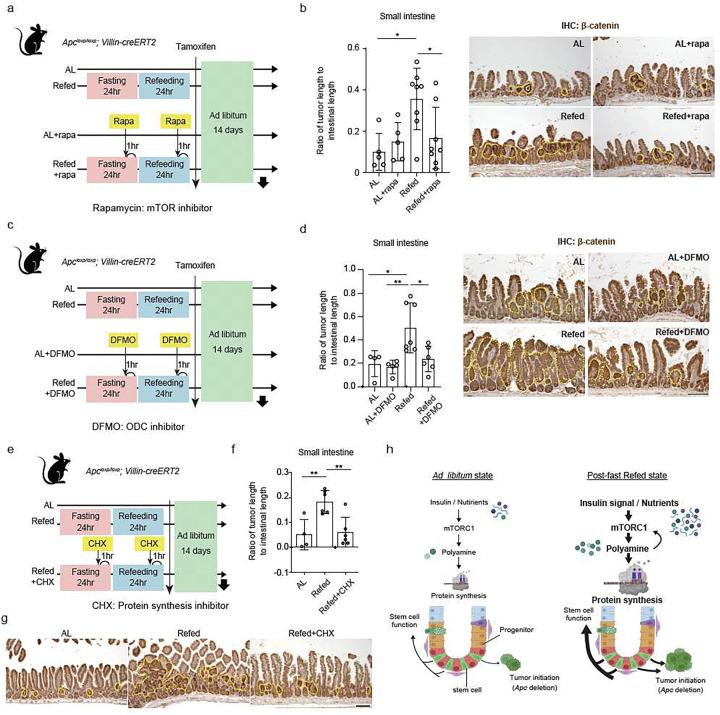
For more than a century, fasting regimens have improved health, lifespan, and tissue regeneration in diverse organisms, including humans. However, how fasting and post-fast refeeding impact adult stem cells and tumour formation has yet to be explored in depth. Here, we demonstrate that post-fast refeeding increases intestinal stem cell (ISC) proliferation and tumour formation: Post-fast refeeding augments the regenerative capacity of Lgr5+ intestinal stem cells (ISCs), and loss of the tumour suppressor Apc in ISCs under post-fast refeeding leads to a higher tumour incidence in the small intestine and colon than in the fasted or ad libitum (AL) fed states. This demonstrates that post-fast refeeding is a distinct state. Mechanistically, we discovered that robust induction of mTORC1 in post-fast-refed ISCs increases protein synthesis via polyamine metabolism to drive these changes, as inhibition of mTORC1, polyamine metabolite production, or protein synthesis abrogates the regenerative or tumourigenic effects of post-fast refeeding. Thus, fast-refeeding cycles must be carefully considered when planning diet-based strategies for regeneration without increasing cancer risk, as post-fast refeeding leads to a burst not only in stem cell-driven regeneration but also in tumourigenicity.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
