

Unraveling signatures of chicken genetic diversity and divergent selection in breed-specific patterns of early myogenesis, nitric oxide metabolism and post-hatch growth
- PMID: 36712856
- PMCID: PMC9874007
- DOI: 10.3389/fgene.2022.1092242
Unraveling signatures of chicken genetic diversity and divergent selection in breed-specific patterns of early myogenesis, nitric oxide metabolism and post-hatch growth
Abstract
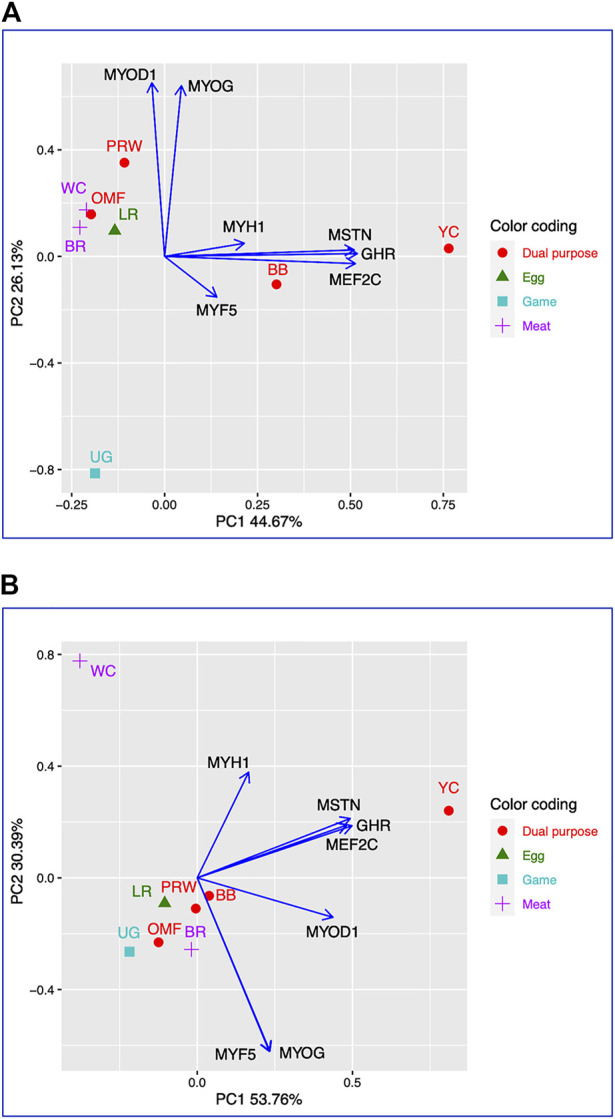
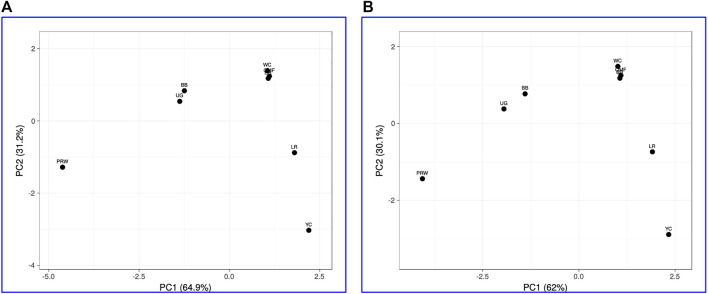
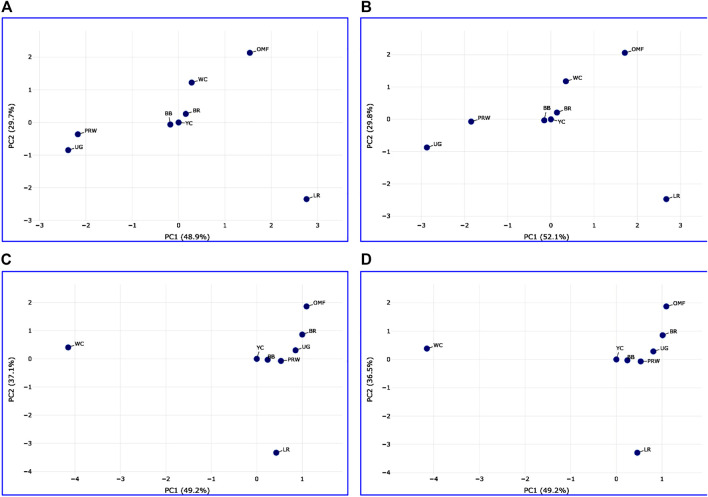
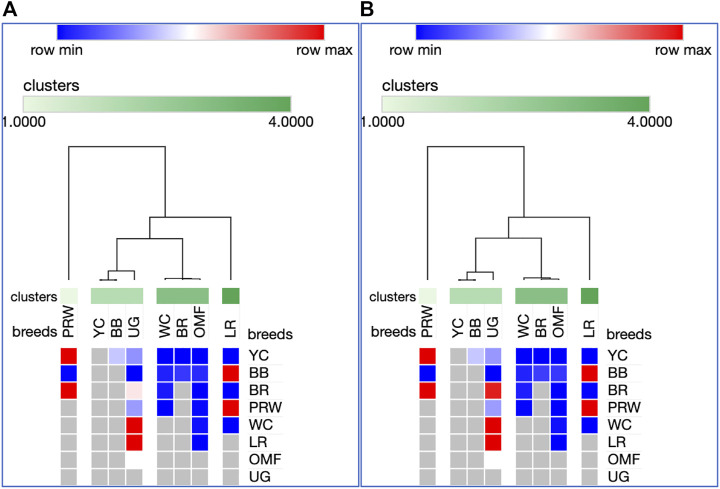
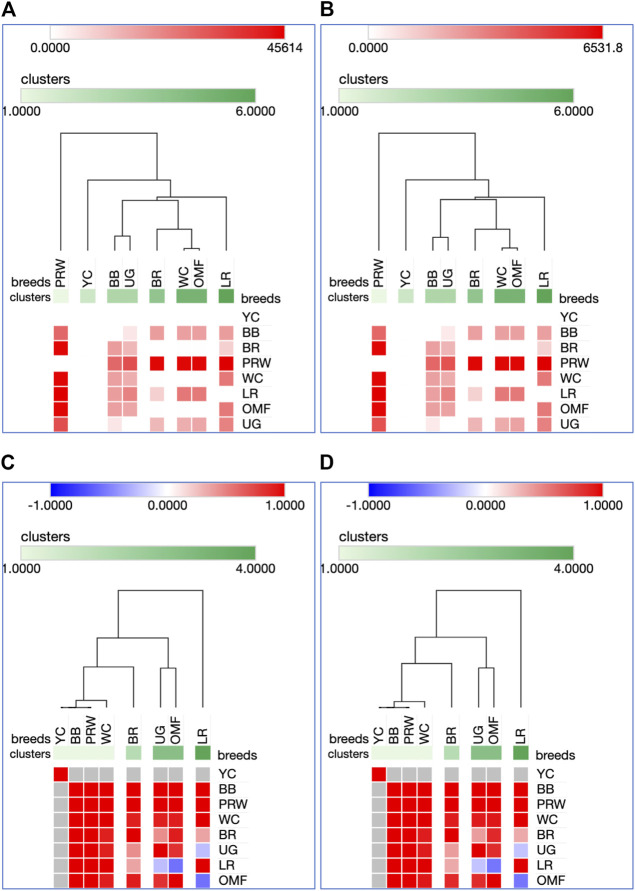
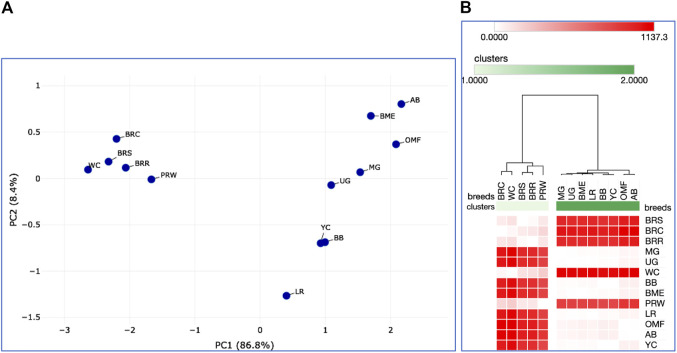
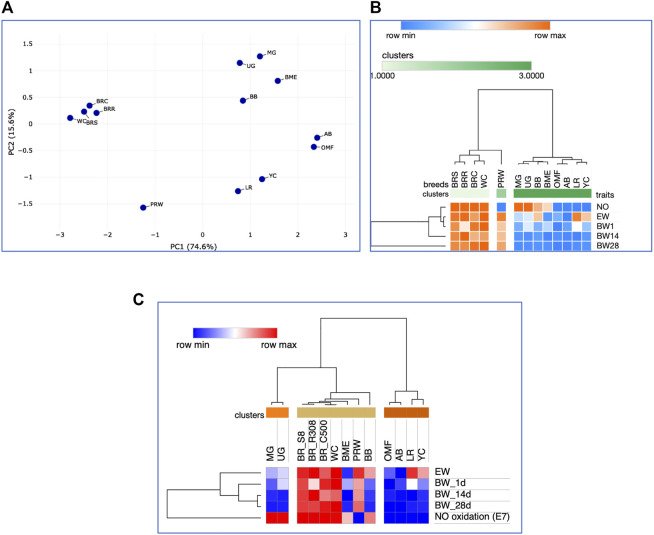
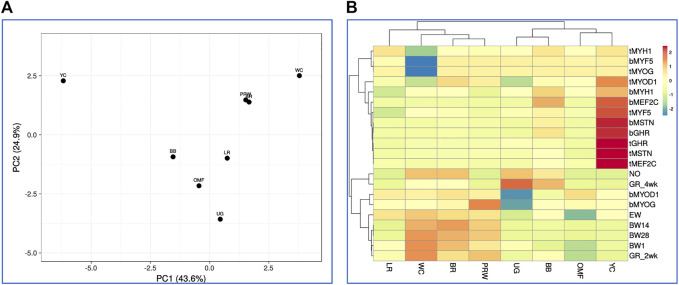
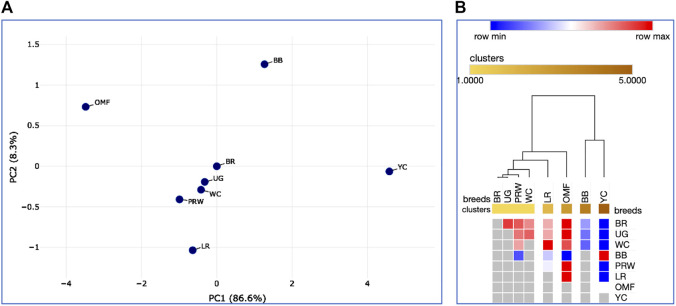
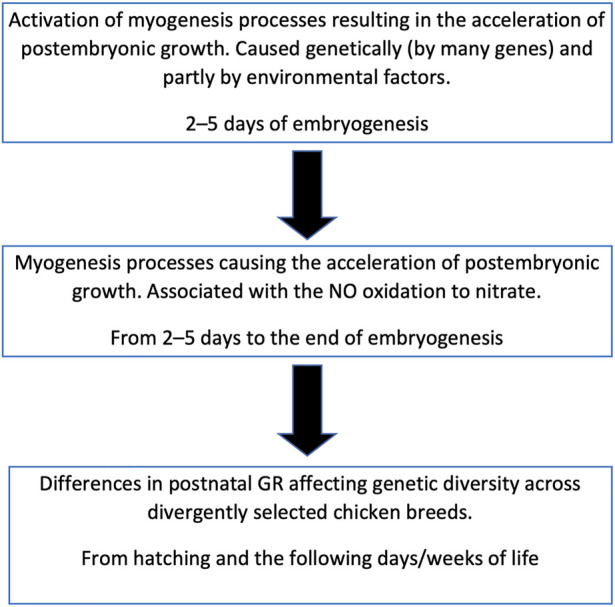
Introduction: Due to long-term domestication, breeding and divergent selection, a vast genetic diversity in poultry currently exists, with various breeds being characterized by unique phenotypic and genetic features. Assuming that differences between chicken breeds divergently selected for economically and culturally important traits manifest as early as possible in development and growth stages, we aimed to explore breed-specific patterns and interrelations of embryo myogenesis, nitric oxide (NO) metabolism and post-hatch growth rate (GR). Methods: These characteristics were explored in eight breeds of different utility types (meat-type, dual purpose, egg-type, game, and fancy) by incubating 70 fertile eggs per breed. To screen the differential expression of seven key myogenesis associated genes (MSTN, GHR, MEF2C, MYOD1, MYOG, MYH1, and MYF5), quantitative real-time PCR was used. Results: We found that myogenesis associated genes expressed in the breast and thigh muscles in a coordinated manner showing breed specificity as a genetic diversity signature among the breeds studied. Notably, coordinated ("accord") expression patterns of MSTN, GHR, and MEFC2 were observed both in the breast and thigh muscles. Also, associated expression vectors were identified for MYOG and MYOD1 in the breast muscles and for MYOG and MYF5 genes in the thigh muscles. Indices of NO oxidation and post-hatch growth were generally concordant with utility types of breeds, with meat-types breeds demonstrating higher NO oxidation levels and greater GR values as compared to egg-type, dual purpose, game and fancy breeds. Discussion: The results of this study suggest that differences in early myogenesis, NO metabolism and post-hatch growth are breed-specific; they appropriately reflect genetic diversity and accurately capture the evolutionary history of divergently selected chicken breeds.
Keywords: breeds; chicken; differential gene expression; divergent selection; early myogenesis; genetic diversity; nitric oxide oxidation; post-hatch growth.
Copyright © 2023 Kochish, Titov, Nikonov, Brazhnik, Vorobyov, Korenyuga, Myasnikova, Dolgorukova, Griffin and Romanov.
Conflict of interest statement
Author EB was employed by the company BIOTROF+ Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Abdelmanova A. S., Dotsev A. V., Romanov M. N., Stanishevskaya O. I., Gladyr E. A., Rodionov A. N., et al. (2021). Unveiling comparative genomic trajectories of selection and key candidate genes in egg-type Russian White and meat-type White Cornish chickens. Biology 10 (9), 876. 10.3390/biology10090876 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous