

Development of an in-vitro high-throughput screening system to identify modulators of genitalia development
- PMID: 36712925
- PMCID: PMC9832959
- DOI: 10.1093/pnasnexus/pgac300
Development of an in-vitro high-throughput screening system to identify modulators of genitalia development
Erratum in
-
Correction to: Volume 2 Issue 1 of PNAS Nexus.PNAS Nexus. 2023 Jan 27;2(1):pgad016. doi: 10.1093/pnasnexus/pgad016. eCollection 2023 Jan. PNAS Nexus. 2023. PMID: 36744020 Free PMC article.
Abstract
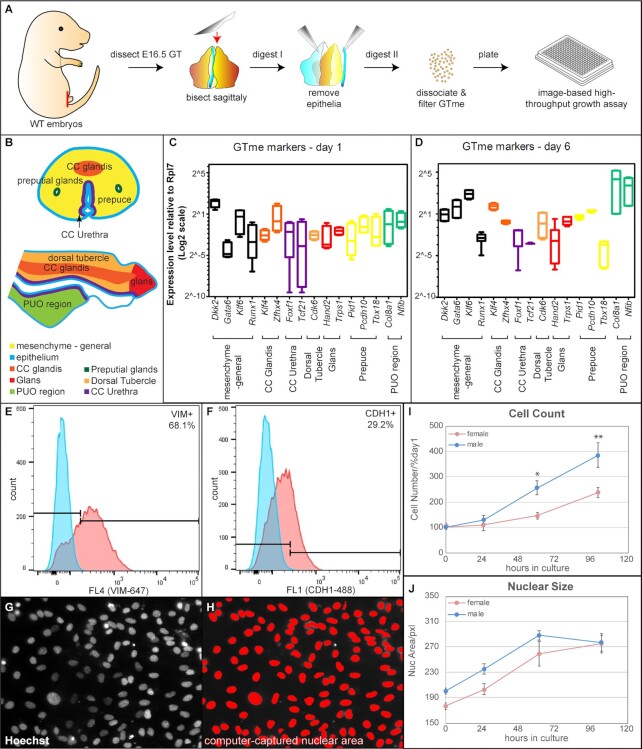
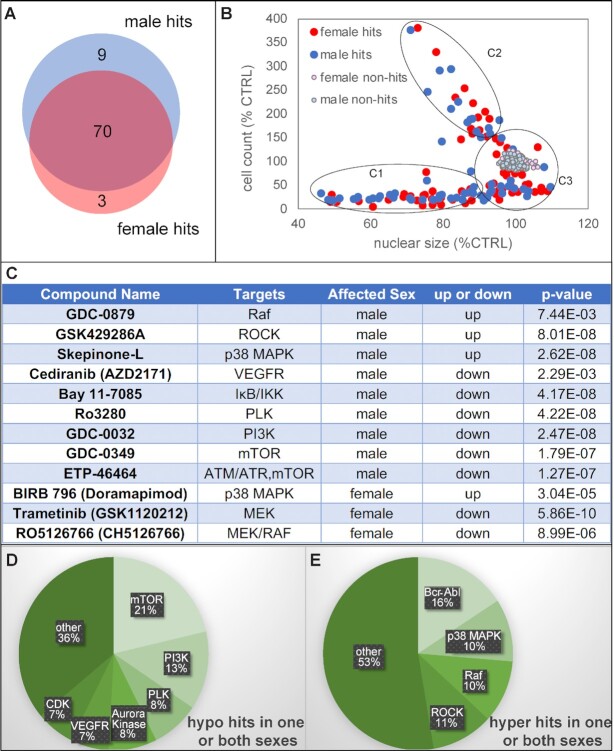
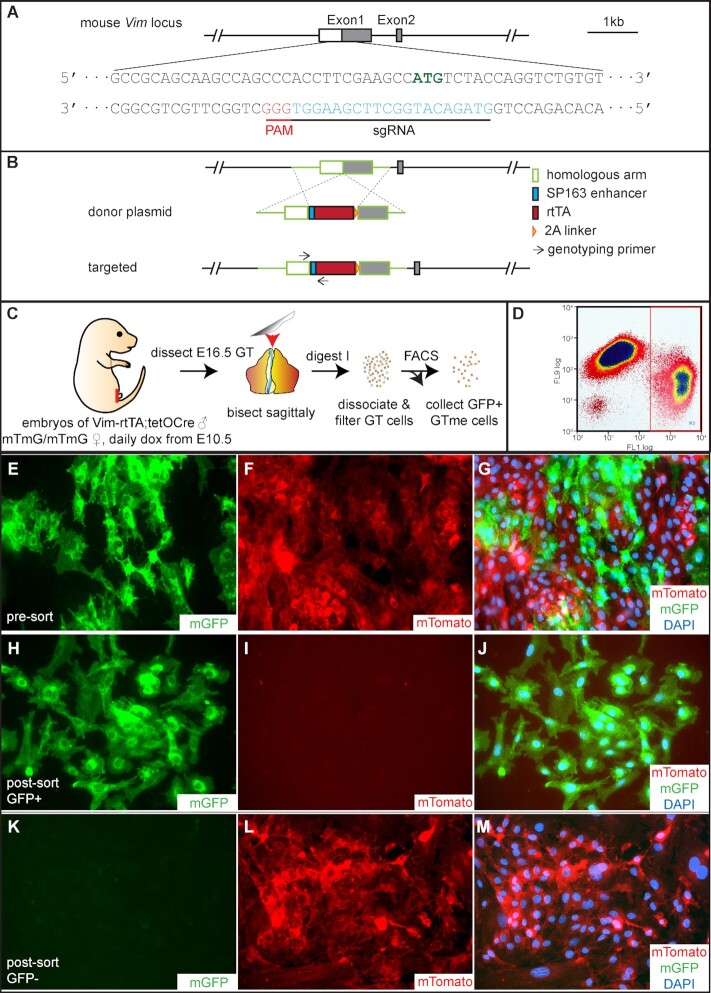
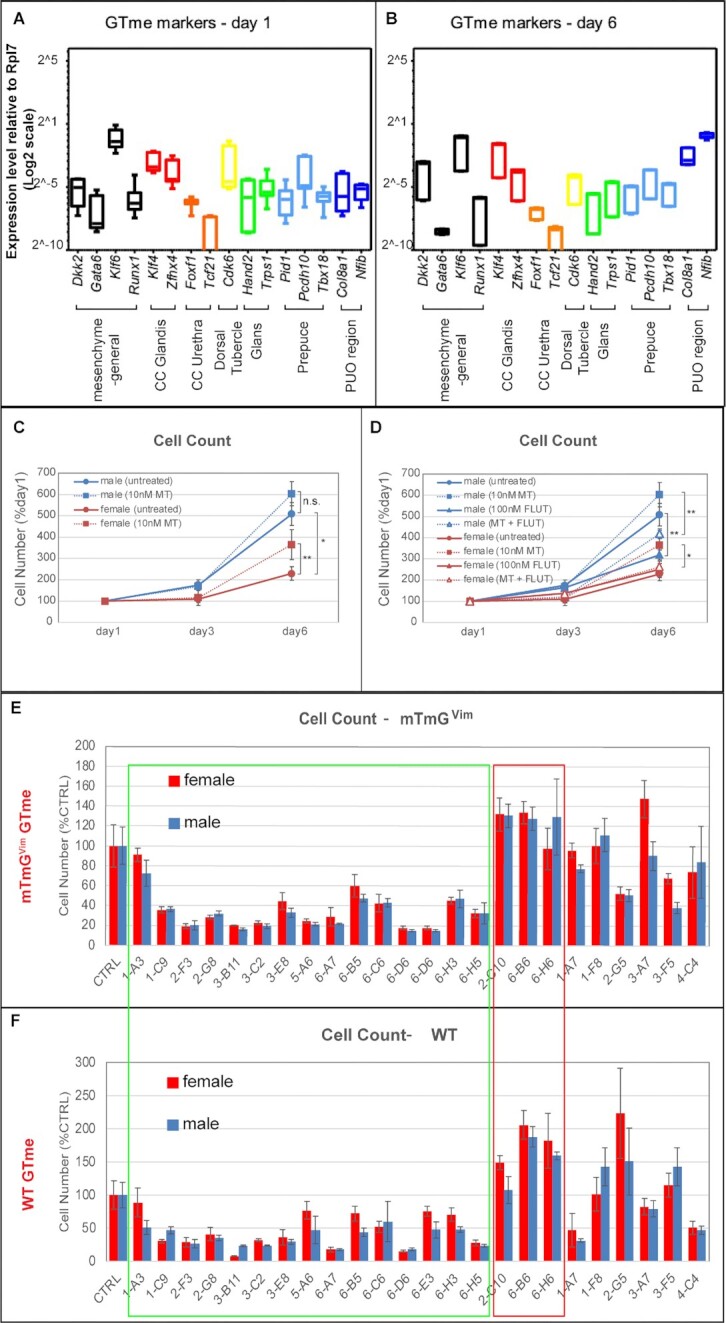
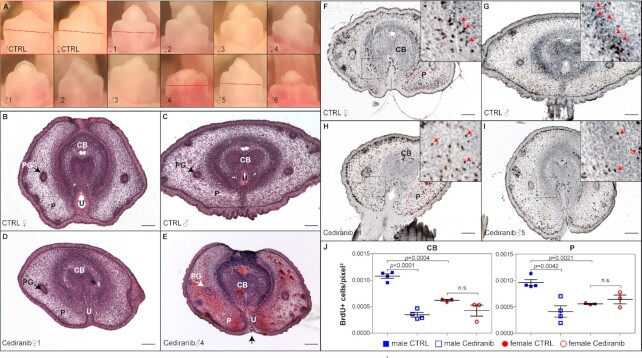
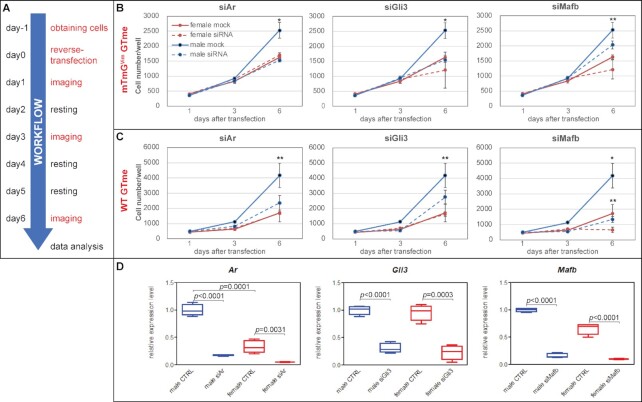
Sexually dimorphic outgrowth and differentiation of the embryonic genital tubercles (GTs) give rise to the penis in males and the clitoris in females. Defects in androgen production or in response to androgen signaling can lead to various congenital penile anomalies in both mice and humans. Due to lack of a high-throughput screening system, identification of crucial regulators of GT sexual differentiation has been slow. To overcome this research barrier, we isolated embryonic GT mesenchymal (GTme) cells to model genitalia growth and differentiation in vitro. Using either a mechanical or fluorescence-activated cell sorting-assisted purification method, GTme cells were isolated and assayed for their proliferation using a microscopy and image analysis system, on a single cell level over time. Male and female GTme cells inherently exhibit different cellular dynamics, consistent with their in-vivo behaviors. This system allows for the rapid quantitative analyses of numerous drug treatments, and enables the discovery of potential genetic modulators of GT morphogenesis on a large scale. Using this system, we completed a 438-compound library screen and identified 82 kinase inhibitor hits. In mice, in-utero exposure to one such candidate kinase inhibitor, Cediranib, resulted in embryos with severe genitalia defects, especially in males. Gene silencing by RNAi was optimized in this system, laying the foundation for future larger-scale genetic screenings. These findings demonstrate the power of this novel high-throughput system to rapidly and successfully identify modulators of genitalia growth and differentiation, expanding the toolbox for the study of functional genomics and environmental factors.
Keywords: VEGFR; external genitalia; genital tubercle mesenchyme; high-throughput screening; proliferation.
The Author(s) 2022. Published by Oxford University Press on behalf of National Academy of Sciences.
Figures
References
-
- Perriton CL, Powles N, Chiang C, Maconochie MK, Cohn MJ. 2002. Sonic hedgehog signaling from the urethral epithelium controls external genital development. Dev Biol. 247:26–46. - PubMed
-
- Haraguchi R, et al. 2001. Unique functions of Sonic hedgehog signaling during external genitalia development. Development. 128:4241–4250. - PubMed
-
- Satoh Y, et al. 2004. Regulation of external genitalia development by concerted actions of FGF ligands and FGF receptors. Anat Embryol (Berl). 208:479–486. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
