

2,3,7,8-Tetrachlorodibenzo-p-dioxin induces multigenerational alterations in the expression of microRNA in the thymus through epigenetic modifications
- PMID: 36712935
- PMCID: PMC9833045
- DOI: 10.1093/pnasnexus/pgac290
2,3,7,8-Tetrachlorodibenzo-p-dioxin induces multigenerational alterations in the expression of microRNA in the thymus through epigenetic modifications
Erratum in
-
Correction to: Volume 2 Issue 1 of PNAS Nexus.PNAS Nexus. 2023 Jan 27;2(1):pgad016. doi: 10.1093/pnasnexus/pgad016. eCollection 2023 Jan. PNAS Nexus. 2023. PMID: 36744020 Free PMC article.
Abstract
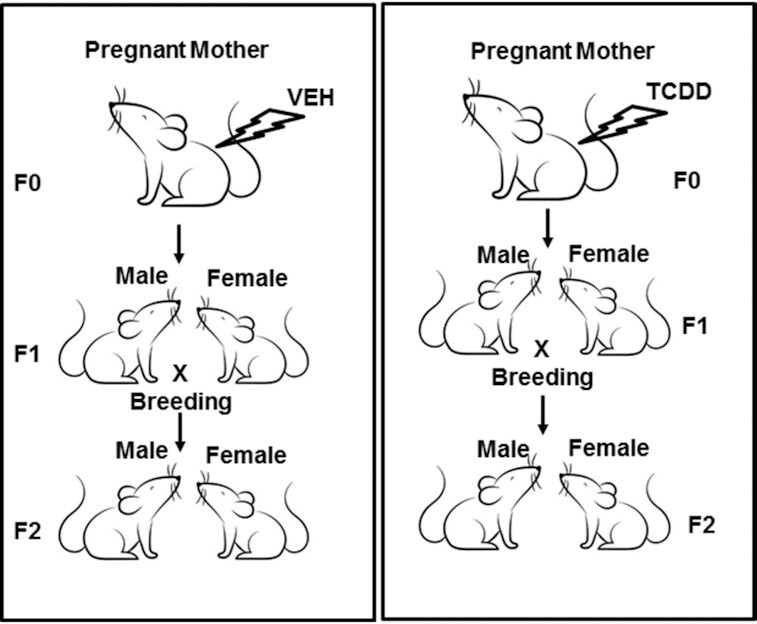
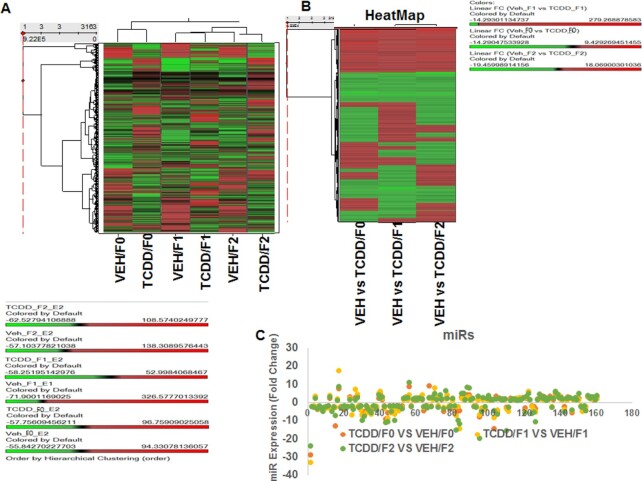
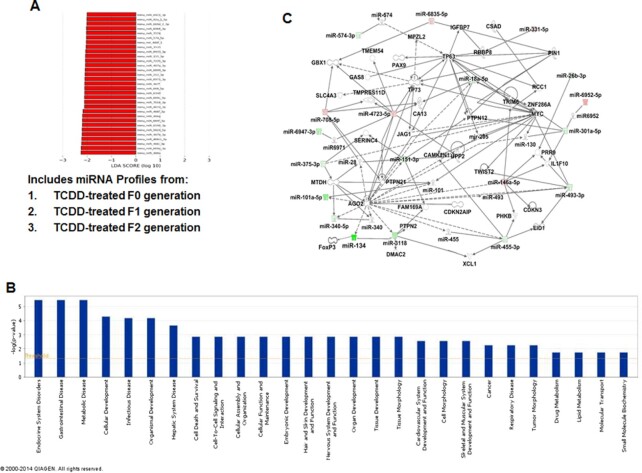
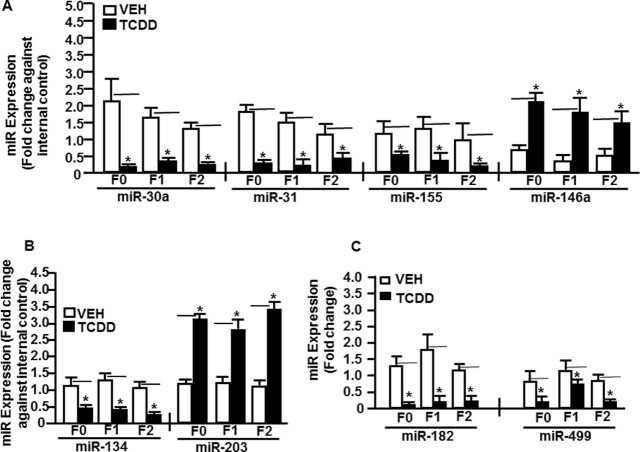
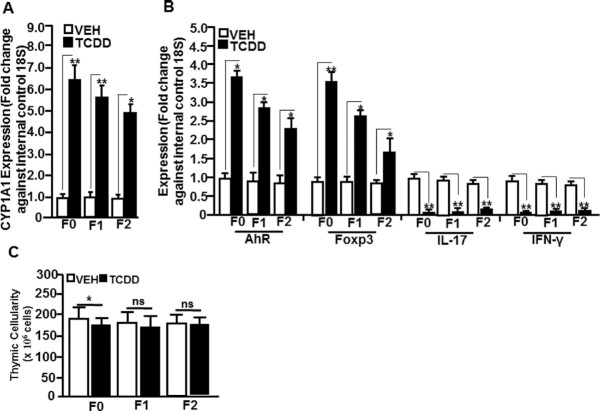
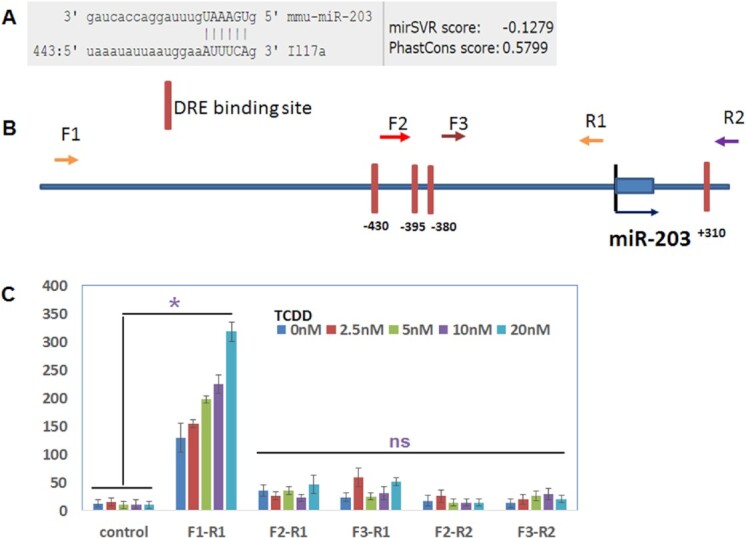
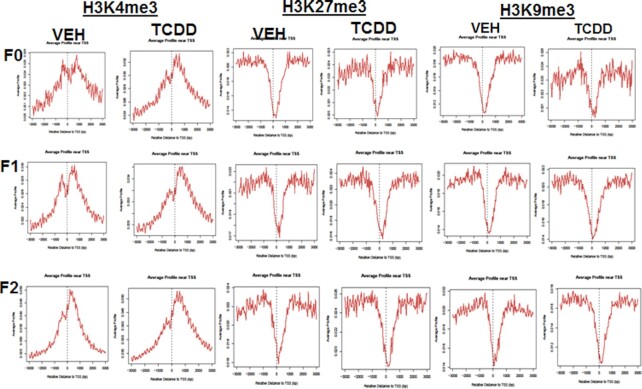
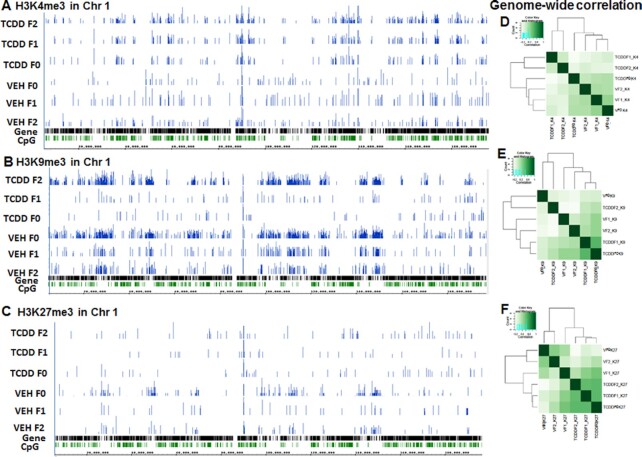
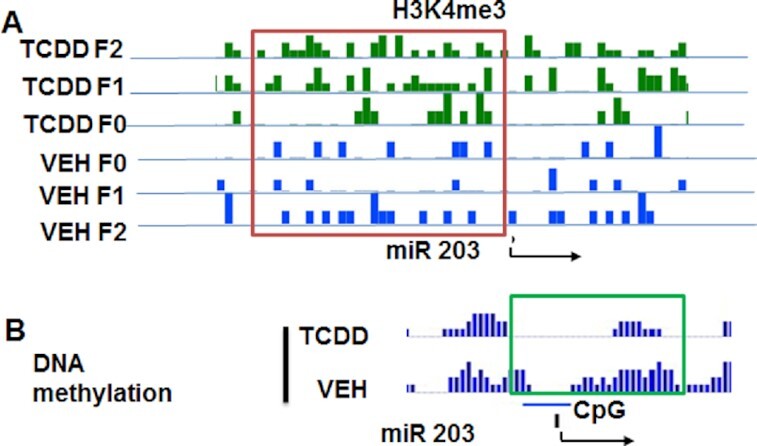
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD), a potent AhR ligand, is an environmental contaminant that is known for mediating toxicity across generations. However, whether TCDD can induce multigenerational changes in the expression of microRNAs (miRs) has not been previously studied. In the current study, we investigated the effect of administration of TCDD in pregnant mice (F0) on gestational day 14, on the expression of miRs in the thymus of F0 and subsequent generations (F1 and F2). Of the 3200 miRs screened, 160 miRs were dysregulated similarly in F0, F1, and F2 generations, while 46 miRs were differentially altered in F0 to F2 generations. Pathway analysis revealed that the changes in miR signature profile mediated by TCDD affected the genes that regulate cell signaling, apoptosis, thymic atrophy, cancer, immunosuppression, and other physiological pathways. A significant number of miRs that showed altered expression exhibited dioxin response elements (DRE) on their promoters. Focusing on one such miR, namely miR-203 that expressed DREs and was induced across F0 to F2 by TCDD, promoter analysis showed that one of the DREs expressed by miR-203 was functional to TCDD-mediated upregulation. Also, the histone methylation status of H3K4me3 in the miR-203 promoter was significantly increased near the transcriptional start site in TCDD-treated thymocytes across F0 to F2 generations. Genome-wide chromatin immunoprecipitation sequencing study suggested that TCDD may cause alterations in histone methylation in certain genes across the three generations. Together, the current study demonstrates that gestational exposure to TCDD can alter the expression of miRs in F0 through direct activation of DREs as well as across F0, F1, and F2 generations through epigenetic pathways.
Keywords: TCDD; epigenetics; histone; microRNA; multigenerational.
© The Author(s) 2022. Published by Oxford University Press on behalf of the National Academy of Sciences.
Figures
References
-
- Gluckman PD, Cutfield W, Hofman P, Hanson MA. 2005. The fetal, neonatal, and infant environments—the long-term consequences for disease risk. Early Hum Dev. 81(1):51–59. - PubMed
-
- Heindel JJ. 2005. The fetal basis of adult disease: role of environmental exposures—introduction. Birth Defects Res A Clin Mol Teratol. 73(3):131–132. - PubMed
-
- Heindel JJ, Levin E. 2005. Developmental origins and environmental influences—introduction. Birth Defects Res A Clin Mol Teratol. 73(7):469. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
