

Extrachromosomal DNA Amplification Contributes to Small Cell Lung Cancer Heterogeneity and Is Associated with Worse Outcomes
- PMID: 36715552
- PMCID: PMC10073312
- DOI: 10.1158/2159-8290.CD-22-0796
Extrachromosomal DNA Amplification Contributes to Small Cell Lung Cancer Heterogeneity and Is Associated with Worse Outcomes
Abstract
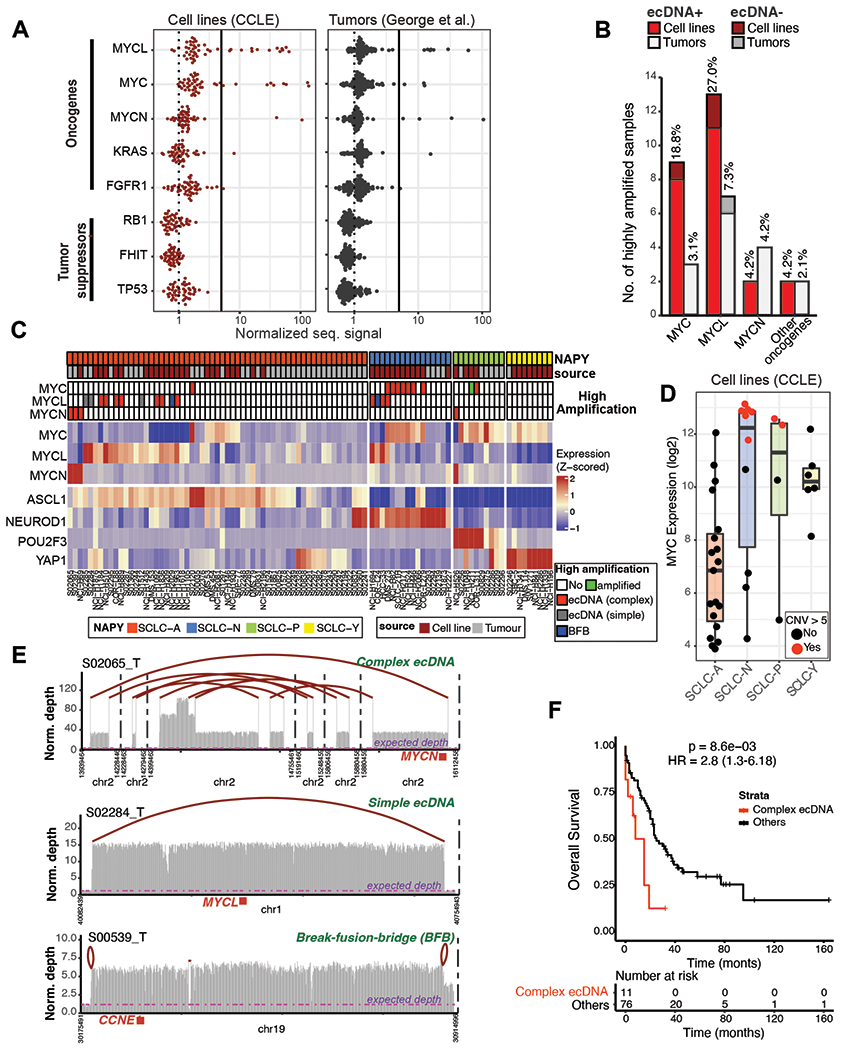
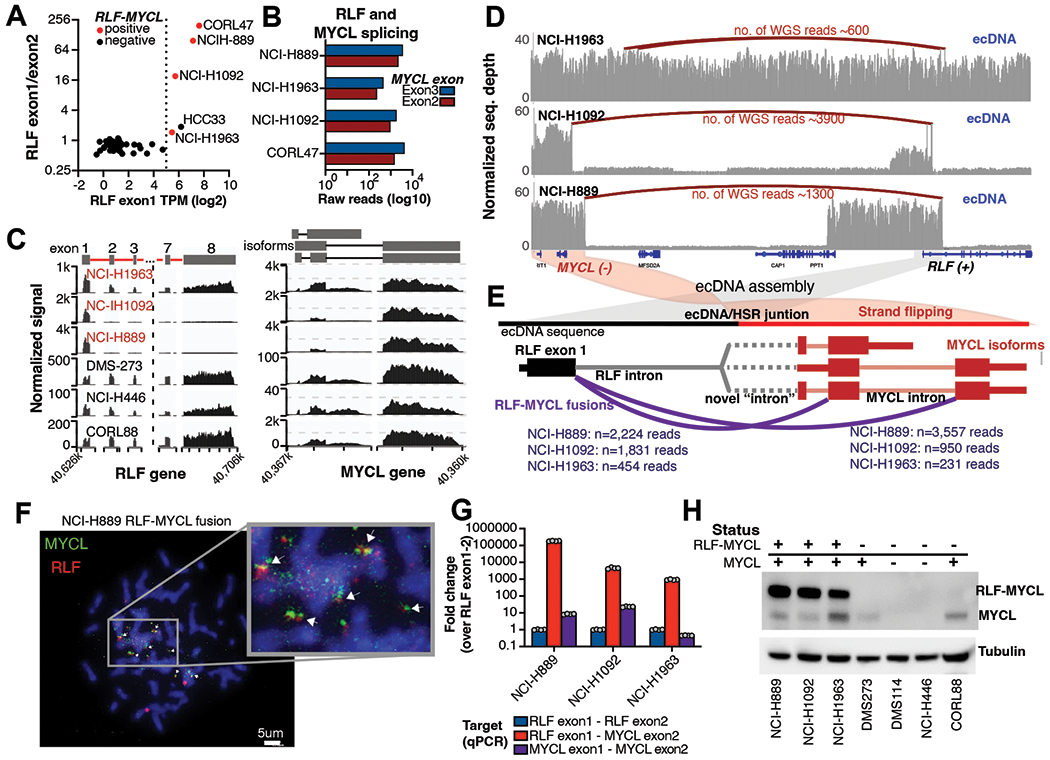
Small-cell lung cancer (SCLC) is an aggressive neuroendocrine lung cancer. Oncogenic MYC amplifications drive SCLC heterogeneity, but the genetic mechanisms of MYC amplification and phenotypic plasticity, characterized by neuroendocrine and nonneuroendocrine cell states, are not known. Here, we integrate whole-genome sequencing, long-range optical mapping, single-cell DNA sequencing, and fluorescence in situ hybridization to find extrachromosomal DNA (ecDNA) as a primary source of SCLC oncogene amplifications and driver fusions. ecDNAs bring to proximity enhancer elements and oncogenes, creating SCLC transcription-amplifying units, driving exceptionally high MYC gene dosage. We demonstrate that cell-free nucleosome profiling can noninvasively detect ecDNA amplifications in plasma, facilitating its genome-wide interrogation in SCLC and other cancers. Altogether, our work provides the first comprehensive map of SCLC ecDNA and describes a new mechanism that governs MYC-driven SCLC heterogeneity. ecDNA-enabled transcriptional flexibility may explain the significantly worse survival outcomes of SCLC harboring complex ecDNA amplifications.
Significance: MYC drives SCLC progression, but the genetic basis of MYC-driven SCLC evolution is unknown. Using SCLC as a paradigm, we report how ecDNA amplifications function as MYC-amplifying units, fostering tumor plasticity and a high degree of tumor heterogeneity. This article is highlighted in the In This Issue feature, p. 799.
©2023 American Association for Cancer Research.
Conflict of interest statement
A. Thomas reports grants to the institution from EMD Serono, Astra Zeneca, Immunomedics, Prolynx, Immunomedics, and Tarveda during the conduct of the study.
Figures




References
-
- Gazdar AF, Carney DN, Nau MM, Minna JD. Characterization of Variant Subclasses of Cell-Lines Derived from Small Cell Lung-Cancer Having Distinctive Biochemical, Morphological, and Growth-Properties. Cancer Res. 1985;45(6):2924–30. - PubMed
-
- Calbo J, van Montfort E, Proost N, van Drunen E, Beverloo HB, Meuwissen R, et al. A Functional Role for Tumor Cell Heterogeneity in a Mouse Model of Small Cell Lung Cancer. Cancer Cell. 2011;19(2):244–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
