

Influence of circadian clocks on adaptive immunity and vaccination responses
- PMID: 36717561
- PMCID: PMC9885059
- DOI: 10.1038/s41467-023-35979-2
Influence of circadian clocks on adaptive immunity and vaccination responses
Abstract
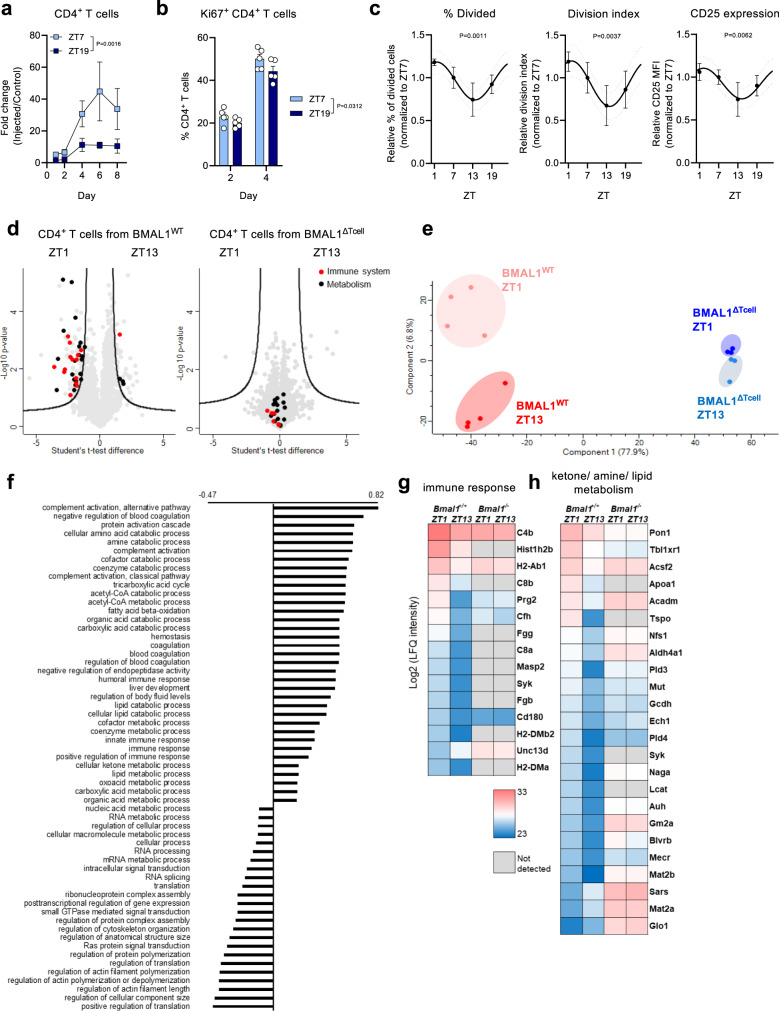
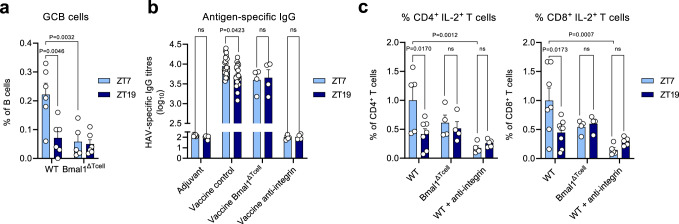
The adaptive immune response is under circadian control, yet, why adaptive immune reactions continue to exhibit circadian changes over long periods of time is unknown. Using a combination of experimental and mathematical modeling approaches, we show here that dendritic cells migrate from the skin to the draining lymph node in a time-of-day-dependent manner, which provides an enhanced likelihood for functional interactions with T cells. Rhythmic expression of TNF in the draining lymph node enhances BMAL1-controlled ICAM-1 expression in high endothelial venules, resulting in lymphocyte infiltration and lymph node expansion. Lymph node cellularity continues to be different for weeks after the initial time-of-day-dependent challenge, which governs the immune response to vaccinations directed against Hepatitis A virus as well as SARS-CoV-2. In this work, we present a mechanistic understanding of the time-of-day dependent development and maintenance of an adaptive immune response, providing a strategy for using time-of-day to optimize vaccination regimes.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
