

Graft-versus-host disease is locally maintained in target tissues by resident progenitor-like T cells
- PMID: 36720219
- PMCID: PMC10182785
- DOI: 10.1016/j.immuni.2023.01.003
Graft-versus-host disease is locally maintained in target tissues by resident progenitor-like T cells
Abstract
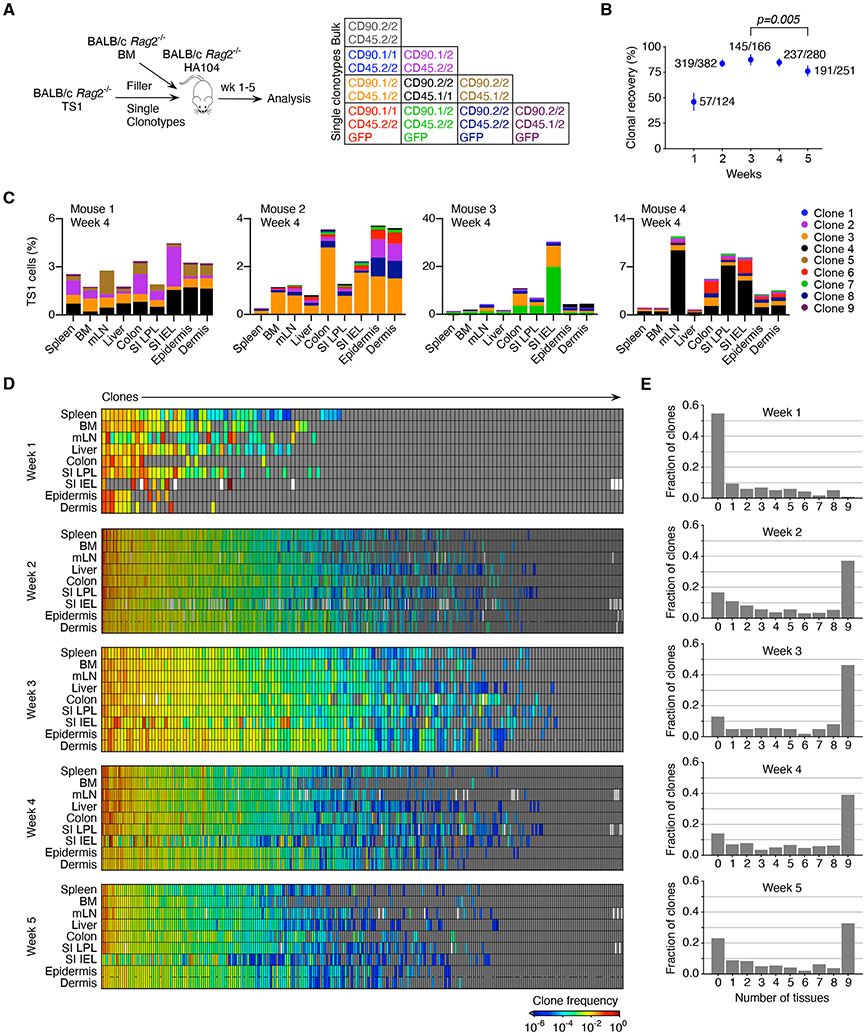
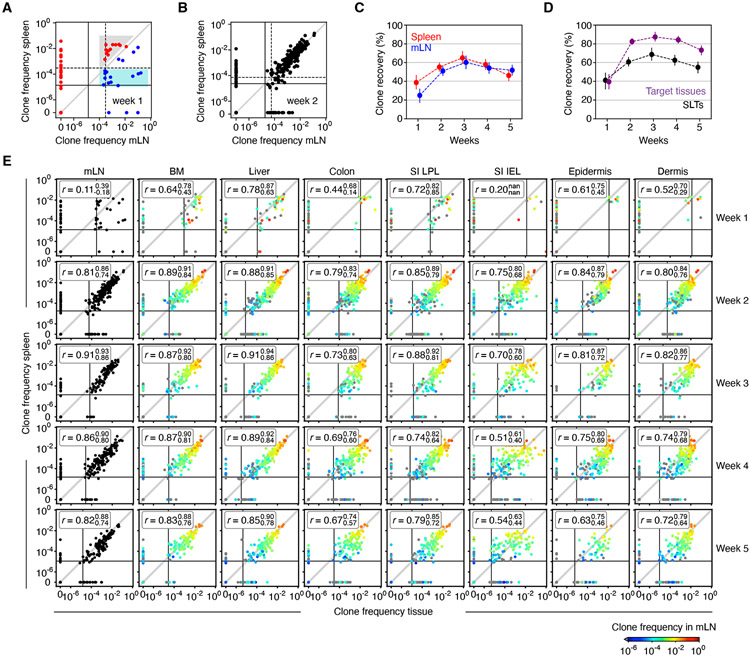
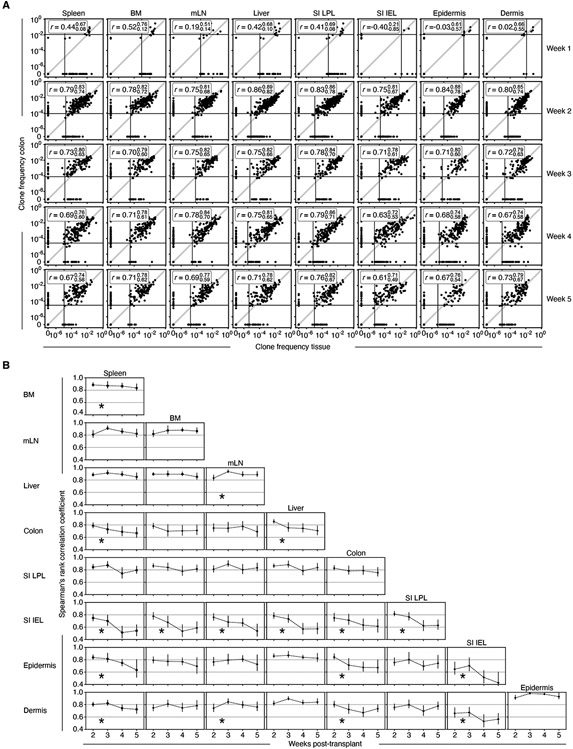
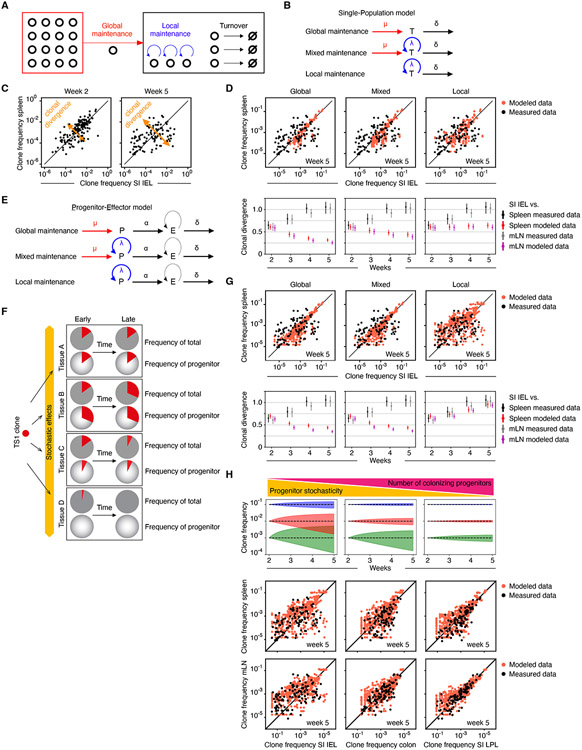
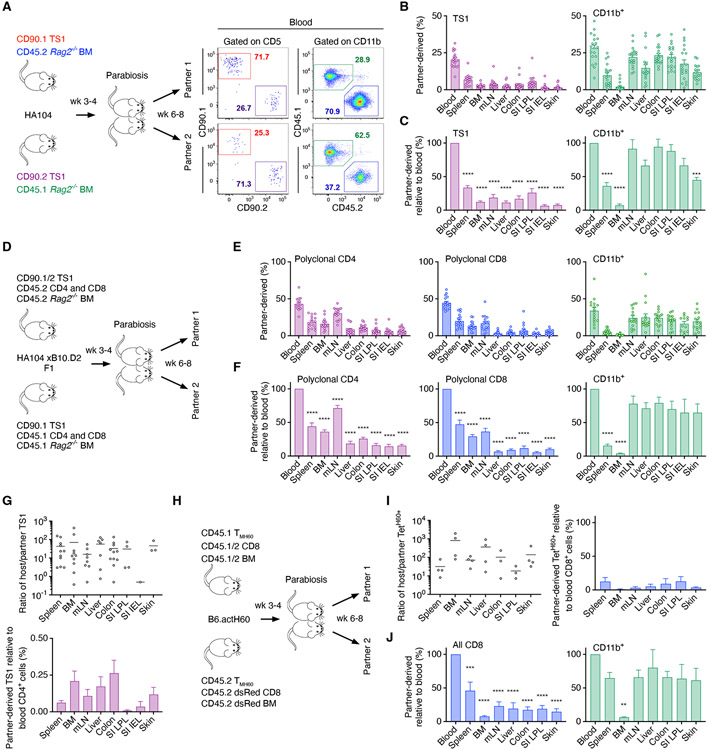
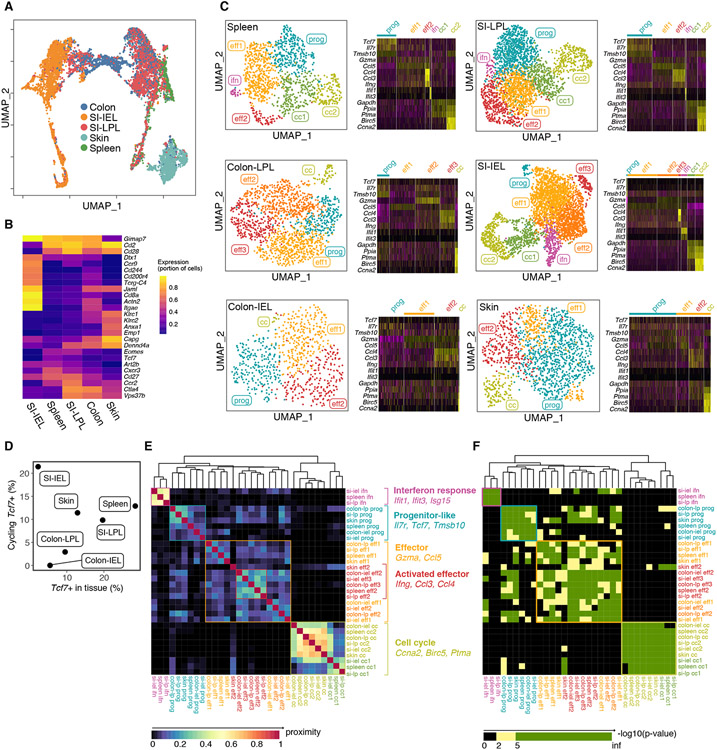
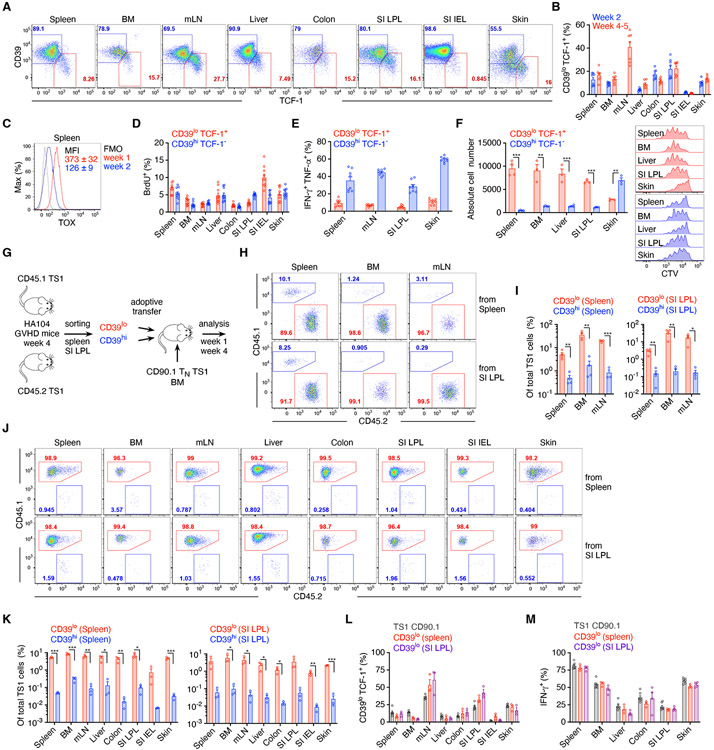
In allogeneic hematopoietic stem cell transplantation, donor αβ T cells attack recipient tissues, causing graft-versus-host disease (GVHD), a major cause of morbidity and mortality. A central question has been how GVHD is sustained despite T cell exhaustion from chronic antigen stimulation. The current model for GVHD holds that disease is maintained through the continued recruitment of alloreactive effectors from blood into affected tissues. Here, we show, using multiple approaches including parabiosis of mice with GVHD, that GVHD is instead primarily maintained locally within diseased tissues. By tracking 1,203 alloreactive T cell clones, we fitted a mathematical model predicting that within each tissue a small number of progenitor T cells maintain a larger effector pool. Consistent with this, we identified a tissue-resident TCF-1+ subpopulation that preferentially engrafted, expanded, and differentiated into effectors upon adoptive transfer. These results suggest that therapies targeting affected tissues and progenitor T cells within them would be effective.
Keywords: T cell exhaustion; TCF-1; TCF-7; Tpex; allogeneic hematopoietic stem cell transplantation; computational modeling; graft-versus-host disease; mouse models; parabiosis; single-cell RNA sequencing.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests W.D.S. is a cofounder, stockholder, and compensated advisor for BlueSphere Bio. He is also a compensated advisor for Orca Bio.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
