

Gut microbiota-derived ursodeoxycholic acid alleviates low birth weight-induced colonic inflammation by enhancing M2 macrophage polarization
- PMID: 36721210
- PMCID: PMC9887892
- DOI: 10.1186/s40168-022-01458-x
Gut microbiota-derived ursodeoxycholic acid alleviates low birth weight-induced colonic inflammation by enhancing M2 macrophage polarization
Abstract
Background: Low birth weight (LBW) is associated with intestinal inflammation and dysbiosis after birth. However, the underlying mechanism remains largely unknown.
Objective: In the present study, we aimed to investigate the metabolism, therapeutic potential, and mechanisms of action of bile acids (BAs) in LBW-induced intestinal inflammation in a piglet model.
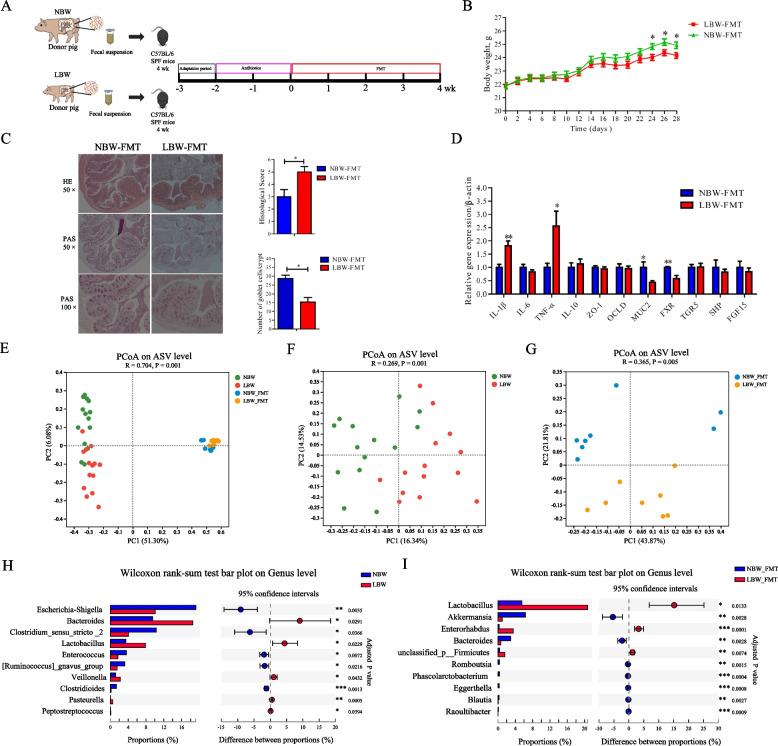
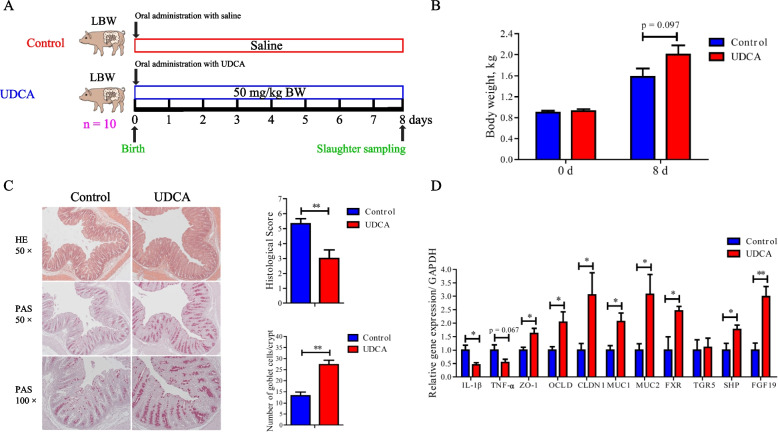
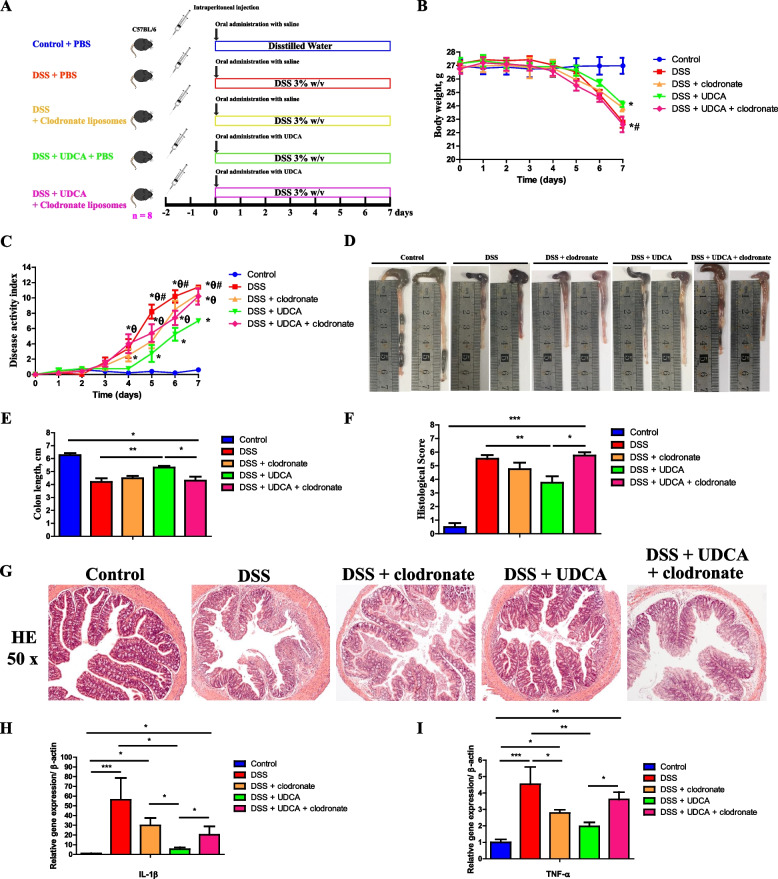
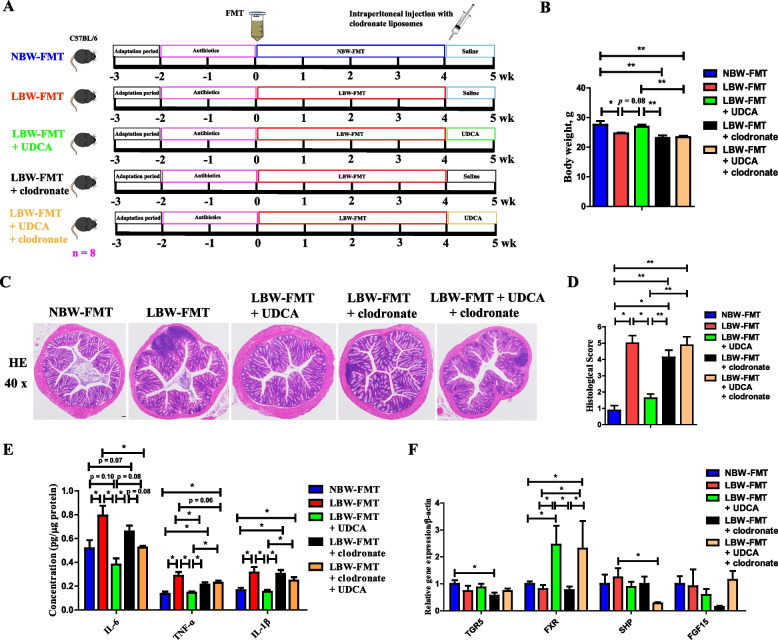
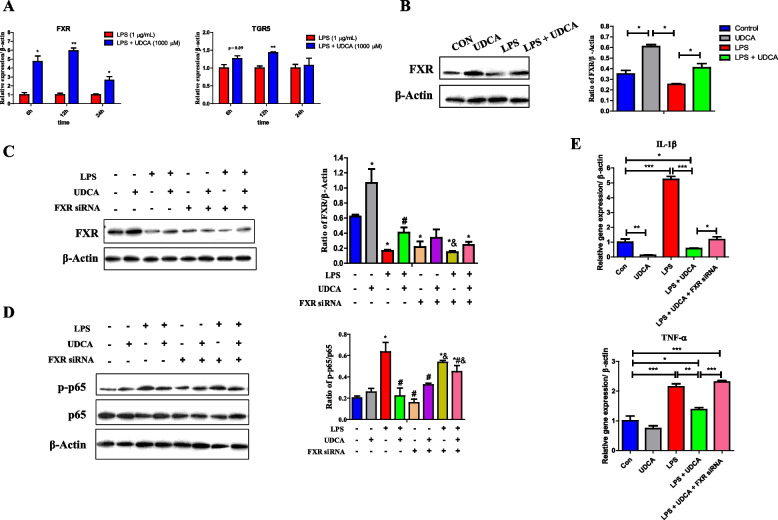
Methods: The fecal microbiome and BA profile between LBW and normal birth weight (NBW) neonatal piglets were compared. Fecal microbiota transplantation (FMT) was employed to further confirm the linkage between microbial BA metabolism and intestinal inflammation. The therapeutic potential of ursodeoxycholic acid (UDCA), a highly differentially abundant BA between LBW and NBW piglets, in alleviating colonic inflammation was evaluated in both LBW piglets, an LBW-FMT mice model, and a DSS-induced colitis mouse model. The underlying cellular and molecular mechanisms by which UDCA suppresses intestinal inflammation were also investigated in both DSS-treated mice and a macrophage cell line. Microbiomes were analyzed by using 16S ribosomal RNA sequencing. Fecal and intestinal BA profiles were measured by using targeted BA metabolomics. Levels of farnesoid X receptor (FXR) were knocked down in J774A.1 cells with small interfering RNAs.
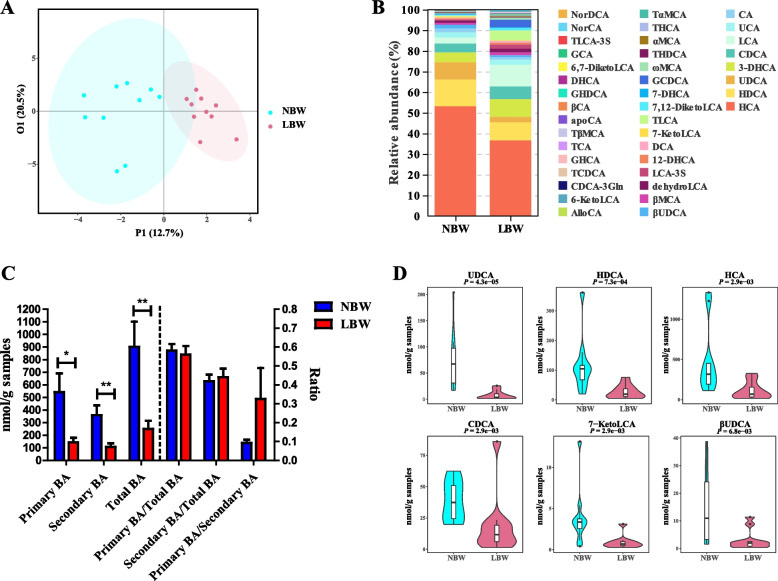
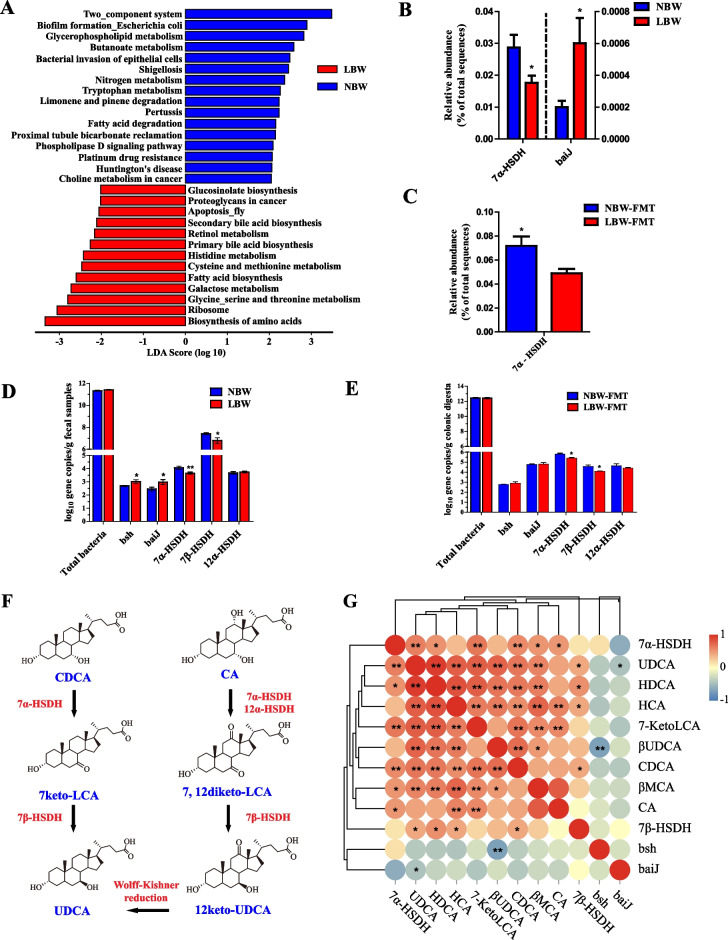
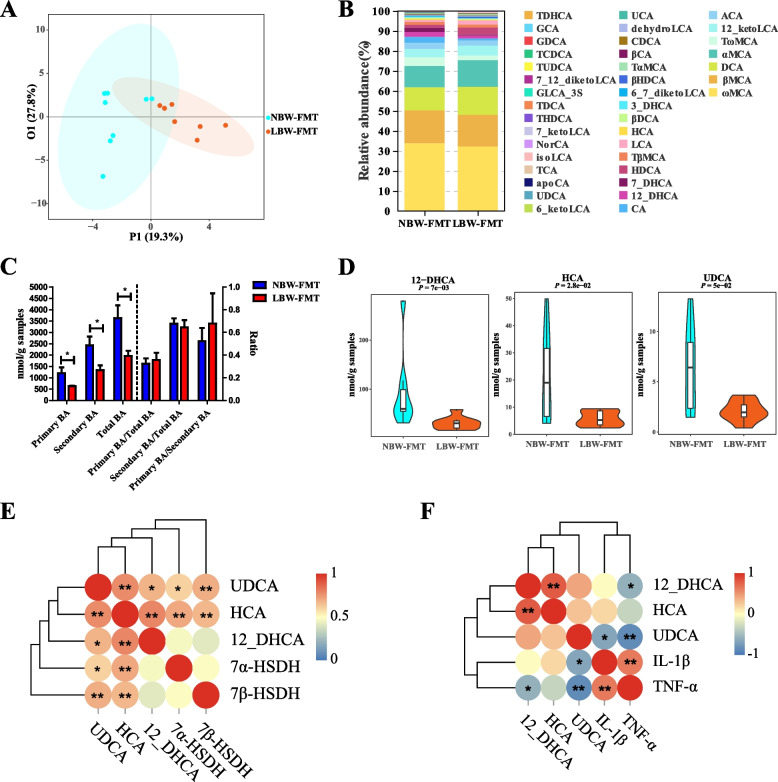
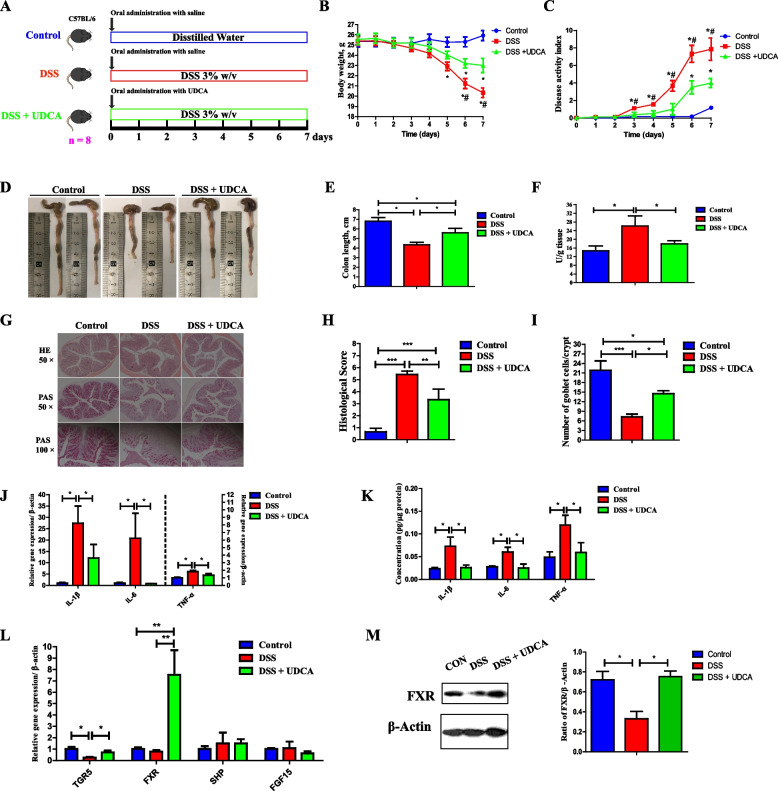
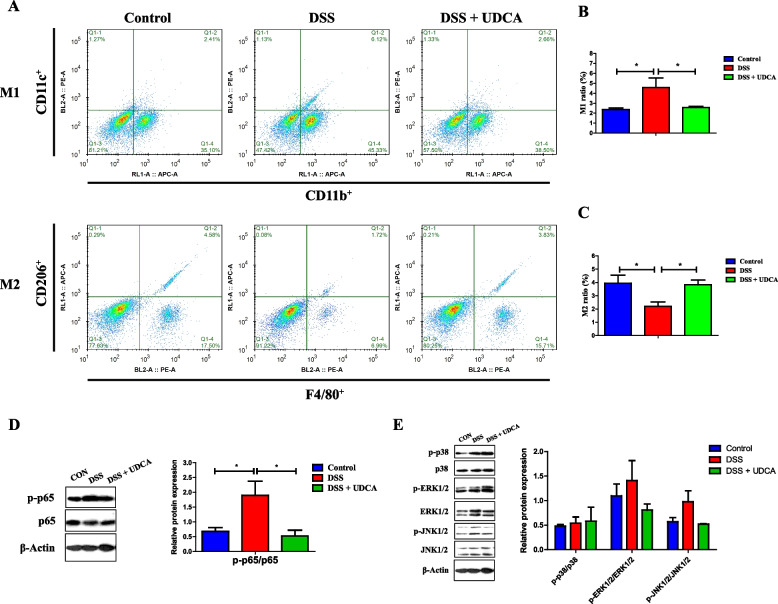
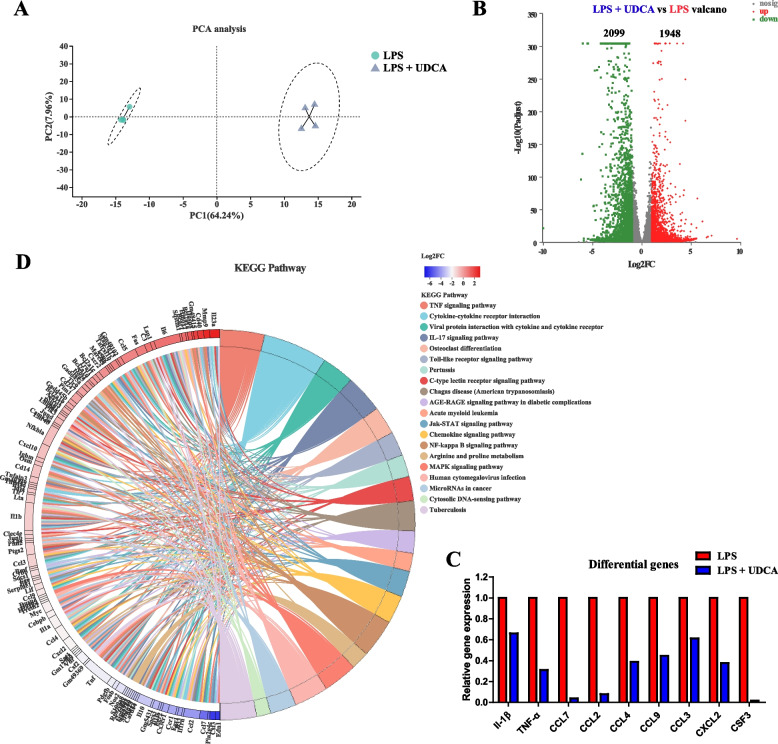
Results: We show a significant difference in both the fecal microbiome and BA profiles between LBW and normal birth weight animals in a piglet model. Transplantation of the microbiota of LBW piglets to antibiotic-treated mice leads to intestinal inflammation. Importantly, oral administration of UDCA, a major BA diminished in the intestinal tract of LBW piglets, markedly alleviates intestinal inflammation in LBW piglets, an LBW-FMT mice model, and a mouse model of colitis by inducing M2 macrophage polarization. Mechanistically, UDCA reduces inflammatory cytokine production by engaging BA receptor FXR while suppressing NF-κB activation in macrophages.
Conclusions: These findings establish a causal relationship between LBW-associated intestinal abnormalities and dysbiosis, suggesting that restoring intestinal health and postnatal maldevelopment of LBW infants may be achieved by targeting intestinal microbiota and BA metabolism. Video Abstract.
Keywords: Bile acids; Immunity; Low birth weight; Macrophage; Microbiome.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- McIntire DD, Bloom SL, Casey BM, Leveno KJ. Birth weight in relation to morbidity and mortality among newborn infants. New Engl J Med. 1999;340(16):1234–1238. - PubMed
-
- Zhu YH, Wang W, Yuan TL, Fu LL, Zhou LA, Lin G, Zhao SH, Zhou HJ, Wu GY, Wang JJ. MicroRNA-29a mediates the impairment of intestinal epithelial integrity induced by intrauterine growth restriction in pig. Am J Physiol-Gastr L. 2017;312(5):G434–G442. - PubMed
-
- Wu G, Bazer FW, Wallace JM, Spencer TE. Board-invited review: intrauterine growth retardation: implications for the animal sciences. J Anim Sci. 2006;84(9):2316–2337. - PubMed
-
- Jain V, Singhal A. Catch up growth in low birth weight infants: striking a healthy balance. Rev Endocr Metab Disord. 2012;13(2):141–147. - PubMed
-
- Guilloteau P, Zabielski R, Hammon HM, Metges CC. Nutritional programming of gastrointestinal tract development. Is the pig a good model for man? Nutr Res Rev. 2010;23(1):4–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
