

Two distinct long-range synaptic complexes promote different aspects of end processing prior to repair of DNA breaks by non-homologous end joining
- PMID: 36724784
- PMCID: PMC9992237
- DOI: 10.1016/j.molcel.2023.01.012
Two distinct long-range synaptic complexes promote different aspects of end processing prior to repair of DNA breaks by non-homologous end joining
Abstract
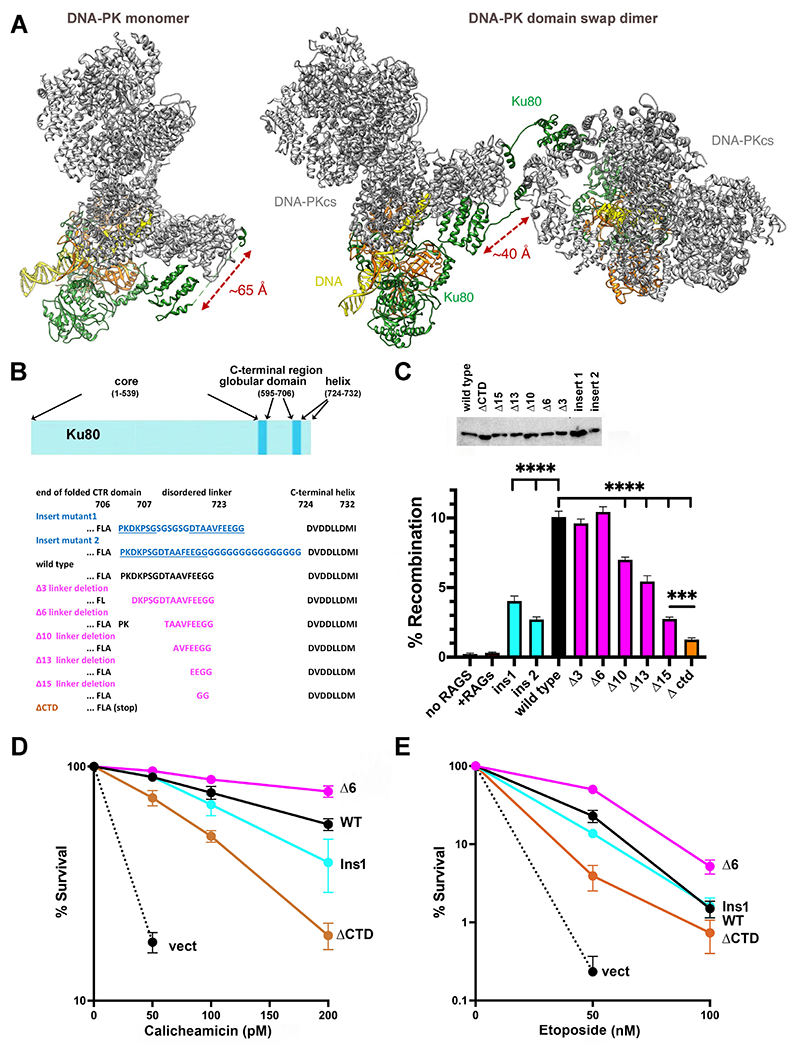
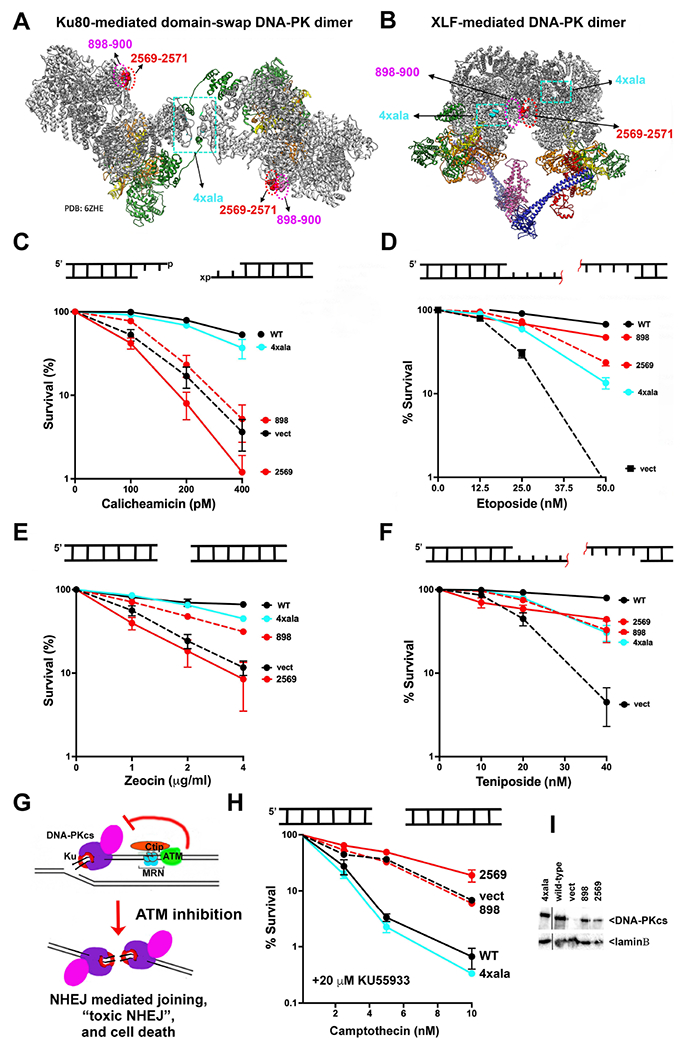
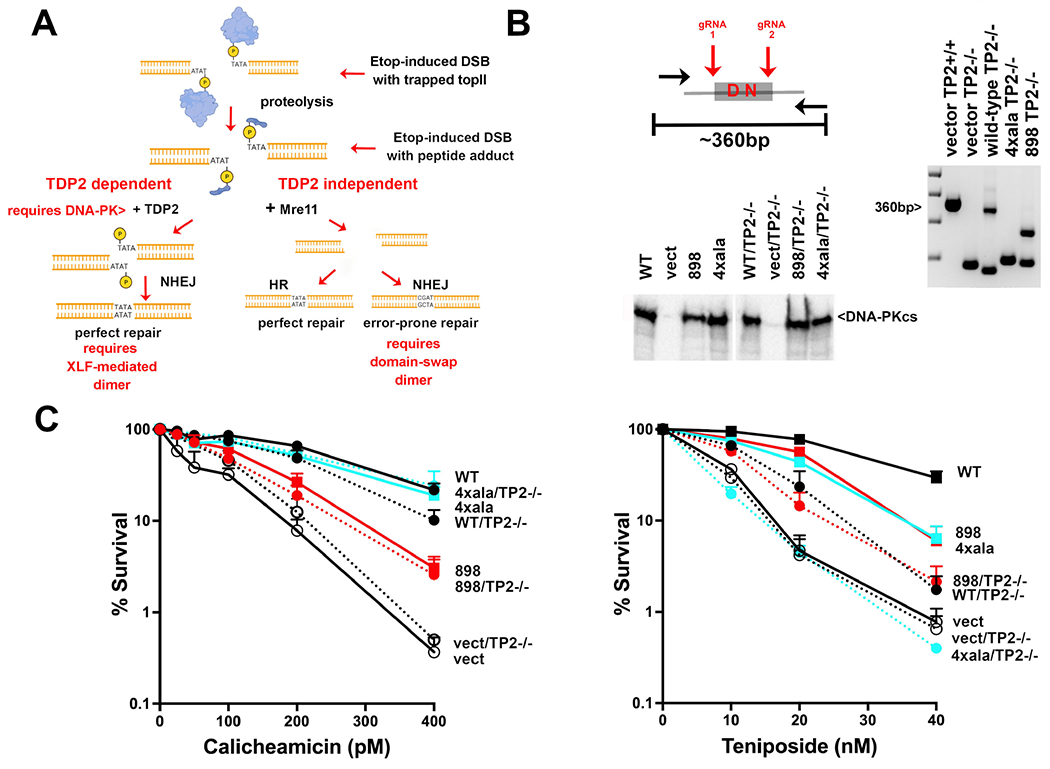
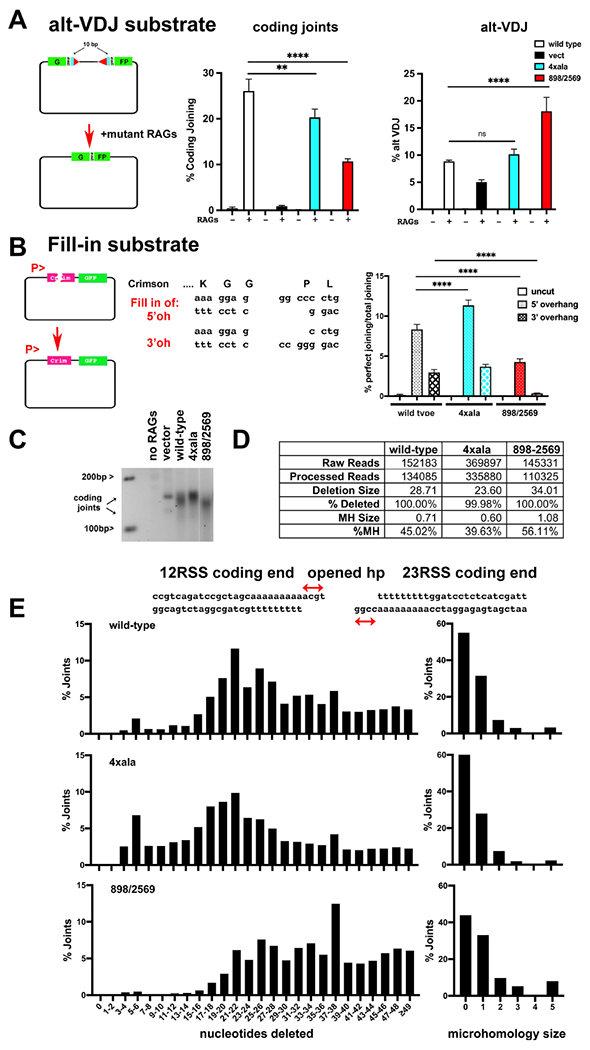
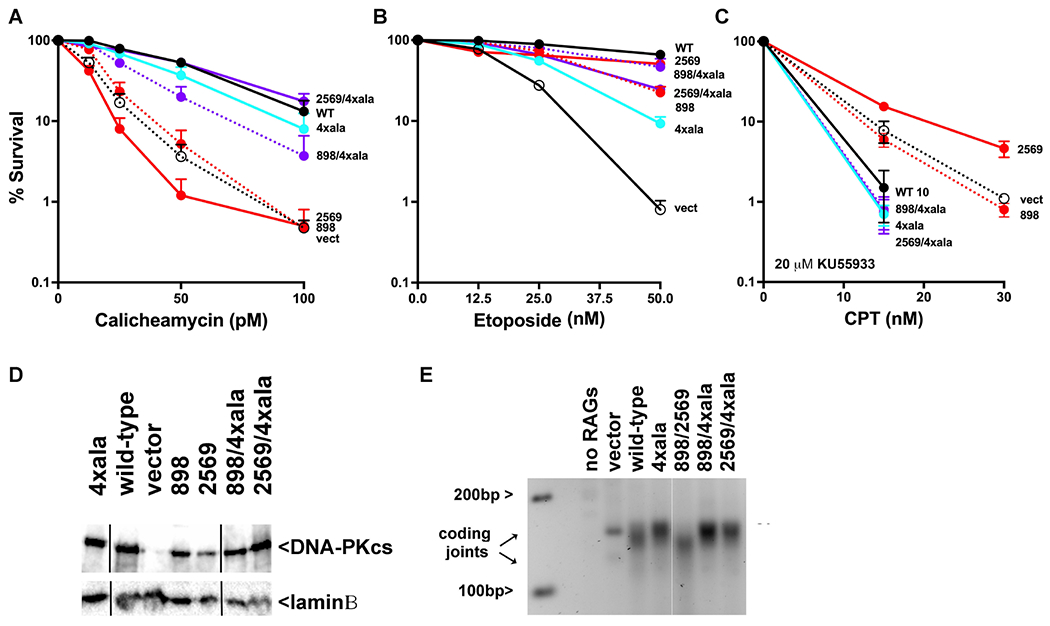
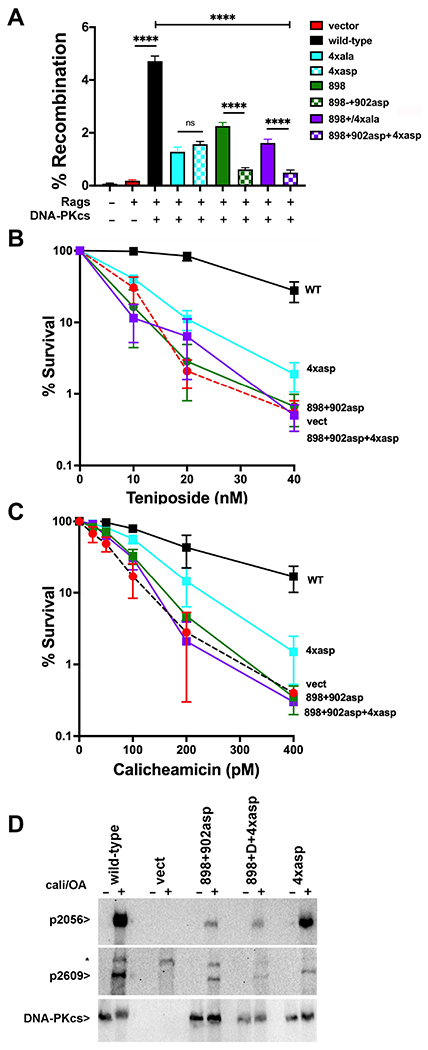
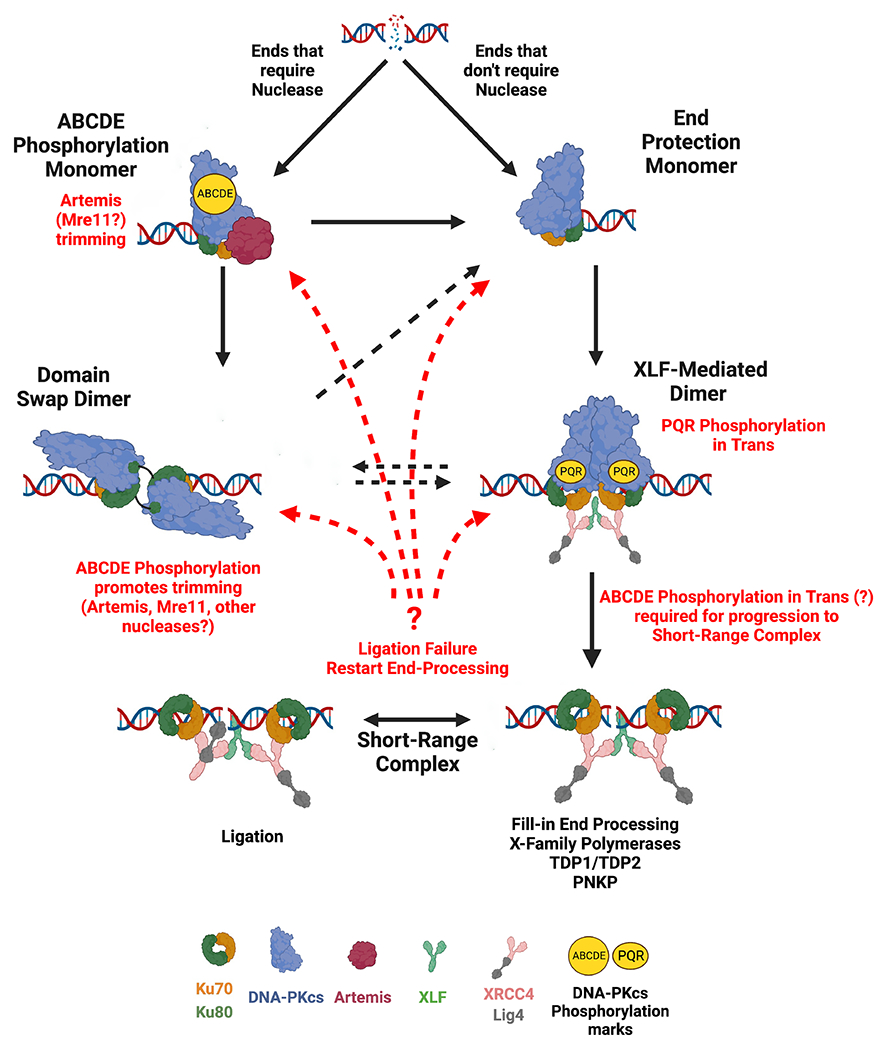
Non-homologous end joining is the major double-strand break repair (DSBR) pathway in mammals. DNA-PK is the hub and organizer of multiple steps in non-homologous end joining (NHEJ). Recent high-resolution structures show how two distinct NHEJ complexes "synapse" two DNA ends. One complex includes a DNA-PK dimer mediated by XLF, whereas a distinct DNA-PK dimer forms via a domain-swap mechanism where the C terminus of Ku80 from one DNA-PK protomer interacts with another DNA-PK protomer in trans. Remarkably, the distance between the two synapsed DNA ends in both dimers is the same (∼115 Å), which matches the distance observed in the initial description of an NHEJ long-range synaptic complex. Here, a mutational strategy is used to demonstrate distinct cellular function(s) of the two dimers: one promoting fill-in end processing, while the other promotes DNA end resection. Thus, the specific DNA-PK dimer formed (which may be impacted by DNA end structure) dictates the mechanism by which ends will be made ligatable.
Keywords: DNA-PK; DNA-PKcs; DNA-dependent protein kinase; Ku; NHEJ; XLF; XRCC4; non-homologous end joining.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Ma Y, Lu H, Schwarz K, and Lieber MR (2005). Repair of double-strand DNA breaks by the human nonhomologous DNA end joining pathway: the iterative processing model. Cell cycle 4, 1193–1200. - PubMed
-
- Waters CA, Strande NT, Pryor JM, Strom CN, Mieczkowski P, Burkhalter MD, Oh S, Qaqish BF, Moore DT, Hendrickson EA, and Ramsden DA (2014). The fidelity of the ligation step determines how ends are resolved during nonhomologous end joining. Nature communications 5, 4286. 10.1038/ncomms5286. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
