

Bi-allelic TTI1 variants cause an autosomal-recessive neurodevelopmental disorder with microcephaly
- PMID: 36724785
- PMCID: PMC10027477
- DOI: 10.1016/j.ajhg.2023.01.006
Bi-allelic TTI1 variants cause an autosomal-recessive neurodevelopmental disorder with microcephaly
Abstract
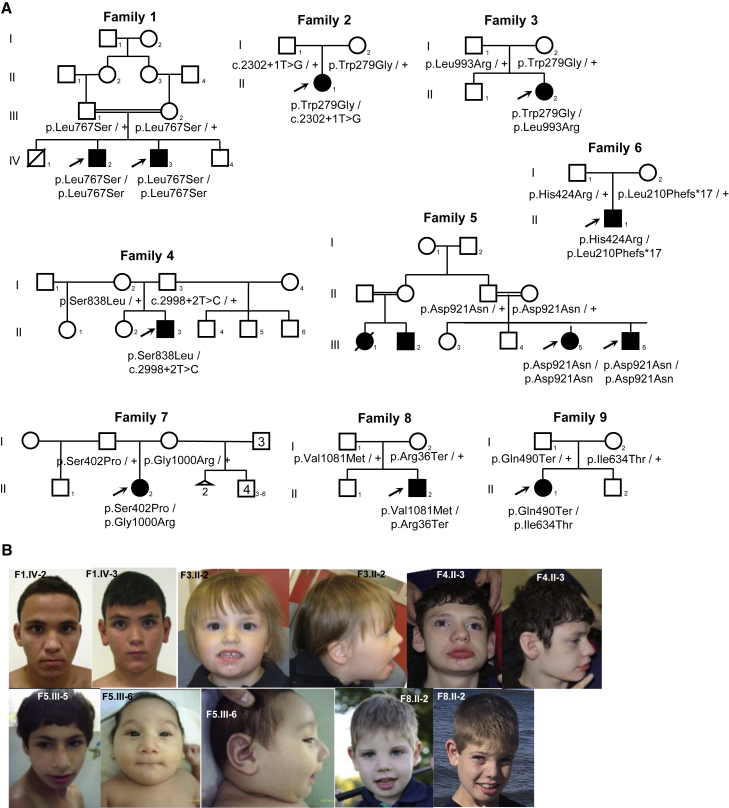
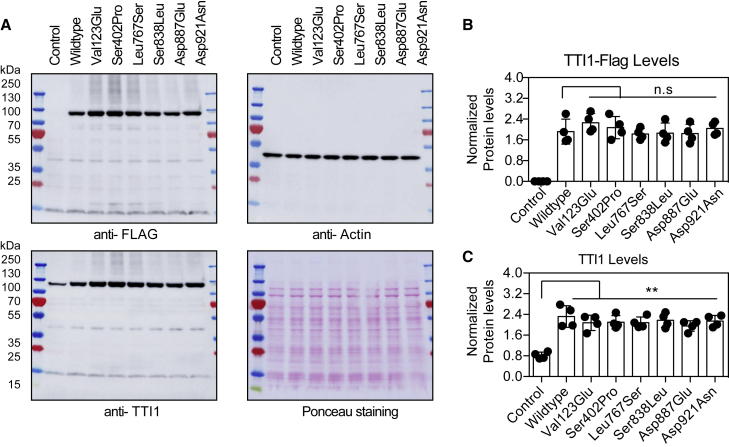
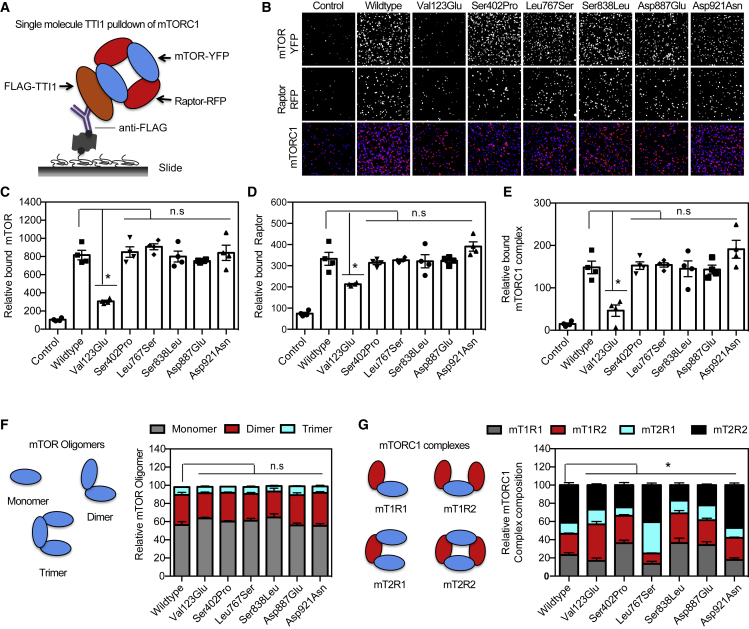
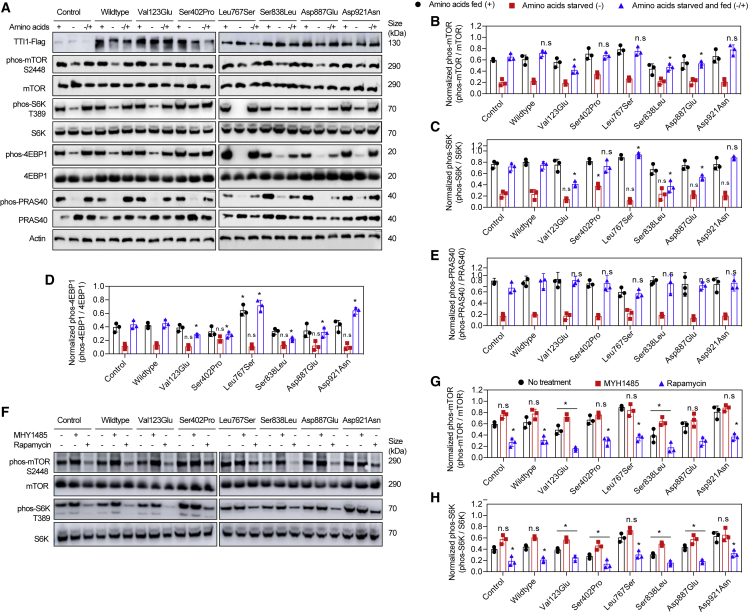
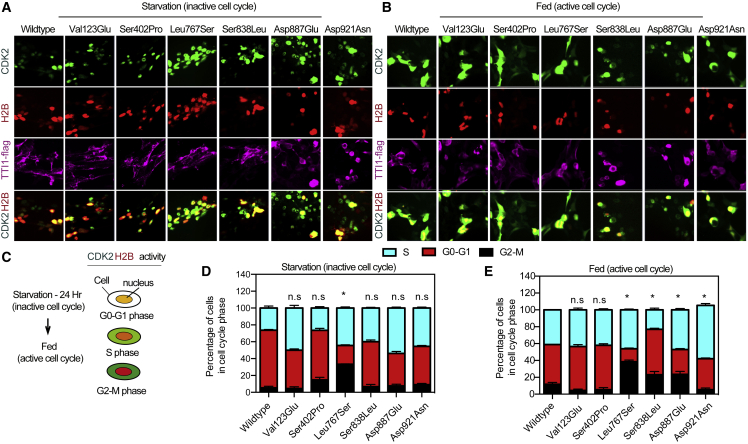
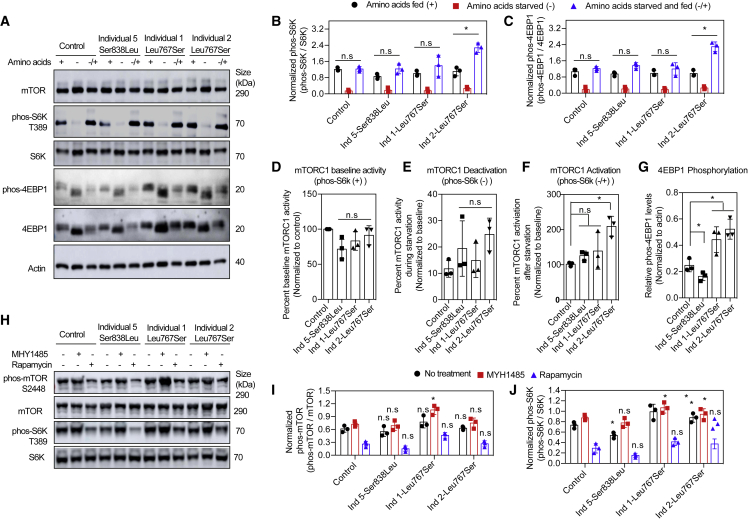
Telomere maintenance 2 (TELO2), Tel2 interacting protein 2 (TTI2), and Tel2 interacting protein 1 (TTI1) are the three components of the conserved Triple T (TTT) complex that modulates activity of phosphatidylinositol 3-kinase-related protein kinases (PIKKs), including mTOR, ATM, and ATR, by regulating the assembly of mTOR complex 1 (mTORC1). The TTT complex is essential for the expression, maturation, and stability of ATM and ATR in response to DNA damage. TELO2- and TTI2-related bi-allelic autosomal-recessive (AR) encephalopathies have been described in individuals with moderate to severe intellectual disability (ID), short stature, postnatal microcephaly, and a movement disorder (in the case of variants within TELO2). We present clinical, genomic, and functional data from 11 individuals in 9 unrelated families with bi-allelic variants in TTI1. All present with ID, and most with microcephaly, short stature, and a movement disorder. Functional studies performed in HEK293T cell lines and fibroblasts and lymphoblastoid cells derived from 4 unrelated individuals showed impairment of the TTT complex and of mTOR pathway activity which is improved by treatment with Rapamycin. Our data delineate a TTI1-related neurodevelopmental disorder and expand the group of disorders related to the TTT complex.
Keywords: TTI1 gene; autosomal recessive; consanguinity; gene; mendelian disorders; microcephaly; neurodevelopment; pathogenic variants.
Copyright © 2023 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.E.A. is a co-founder and CEO of Medigenome, Swiss Institute of Genomic Medicine, and serves in the Scientific Advisory Board of the “Imagine” Institute in Paris. The Department of Medical and Human Genetics at Baylor College of Medicine receives revenue from clinical genetic testing conducted at Baylor Genetics Laboratories. J.R.L. has stock ownership in 23andMe; is a paid consultant for Regeneron Genetics Center; and is a co-inventor on multiple United States and European patents related to molecular diagnostics for inherited neuropathies, eye diseases, genomic disorders, and bacterial genomic fingerprinting, and serves on the Scientific Advisory Board of Baylor Genomics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
