

Cell type specific IL-27p28 (IL-30) deletion in mice uncovers an unexpected regulatory function of IL-30 in autoimmune inflammation
- PMID: 36725904
- PMCID: PMC9892501
- DOI: 10.1038/s41598-023-27413-w
Cell type specific IL-27p28 (IL-30) deletion in mice uncovers an unexpected regulatory function of IL-30 in autoimmune inflammation
Abstract
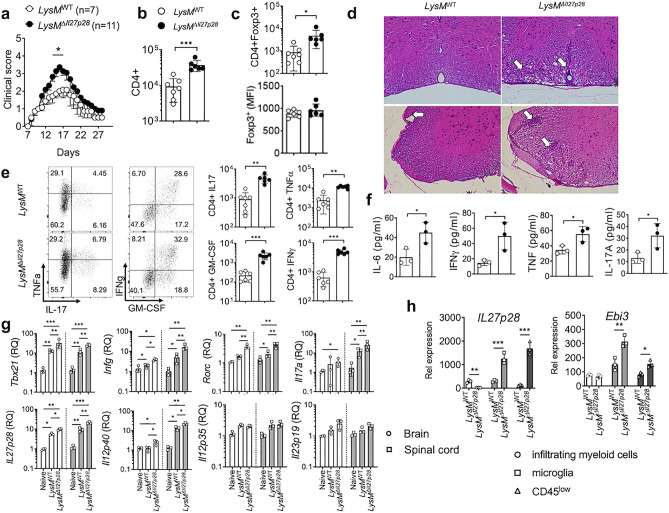
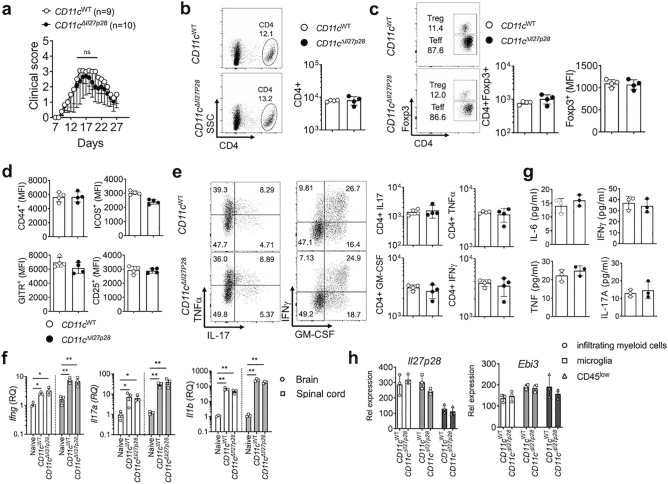
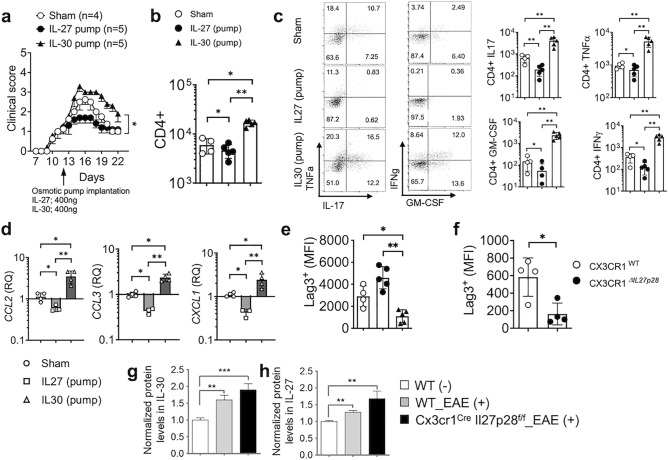
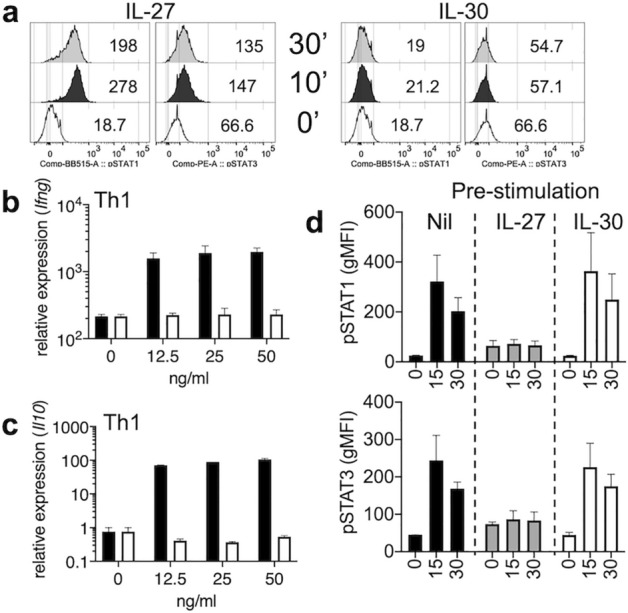
IL-27 is an IL-12 family cytokine with immune regulatory properties, capable of modulating inflammatory responses, including autoimmunity. While extensive studies investigated the major target cells of IL-27 mediating its functions, the source of IL-27 especially during tissue specific autoimmune inflammation has not formally been examined. IL-27p28 subunit, also known as IL-30, was initially discovered as an IL-27-specific subunit, and it has thus been deemed as a surrogate marker to denote IL-27 expression. However, IL-30 can be secreted independently of Ebi3, a subunit that forms bioactive IL-27 with IL-30. Moreover, IL-30 itself may act as a negative regulator antagonizing IL-27. In this study, we exploited various cell type specific IL-30-deficient mouse models and examined the source of IL-30 in a T cell mediated autoimmune neuroinflammation. We found that IL-30 expressed by infiltrating and CNS resident APC subsets, infiltrating myeloid cells and microglia, is central in limiting the inflammation. However, dendritic cell-derived IL-30 was dispensable for the disease development. Unexpectedly, in cell type specific IL-30 deficient mice that develop severe EAE, IL-30 expression in the remaining wild-type APC subsets is disproportionately increased, suggesting that increased endogenous IL-30 production may be involved in the severe pathogenesis. In support, systemic recombinant IL-30 administration exacerbates EAE severity. Our results demonstrate that dysregulated endogenous IL-30 expression may interfere with immune regulatory functions of IL-27, promoting encephalitogenic inflammation in vivo.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Yoshimura T, et al. Two-sided roles of IL-27: Induction of Th1 differentiation on naive CD4+ T cells versus suppression of proinflammatory cytokine production including IL-23-induced IL-17 on activated CD4+ T cells partially through STAT3-dependent mechanism. J. Immunol. 2006;177:5377–5385. doi: 10.4049/jimmunol.177.8.5377. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
