

Associations of psychiatric disease and ageing with FKBP5 expression converge on superficial layer neurons of the neocortex
- PMID: 36729133
- PMCID: PMC10020280
- DOI: 10.1007/s00401-023-02541-9
Associations of psychiatric disease and ageing with FKBP5 expression converge on superficial layer neurons of the neocortex
Abstract
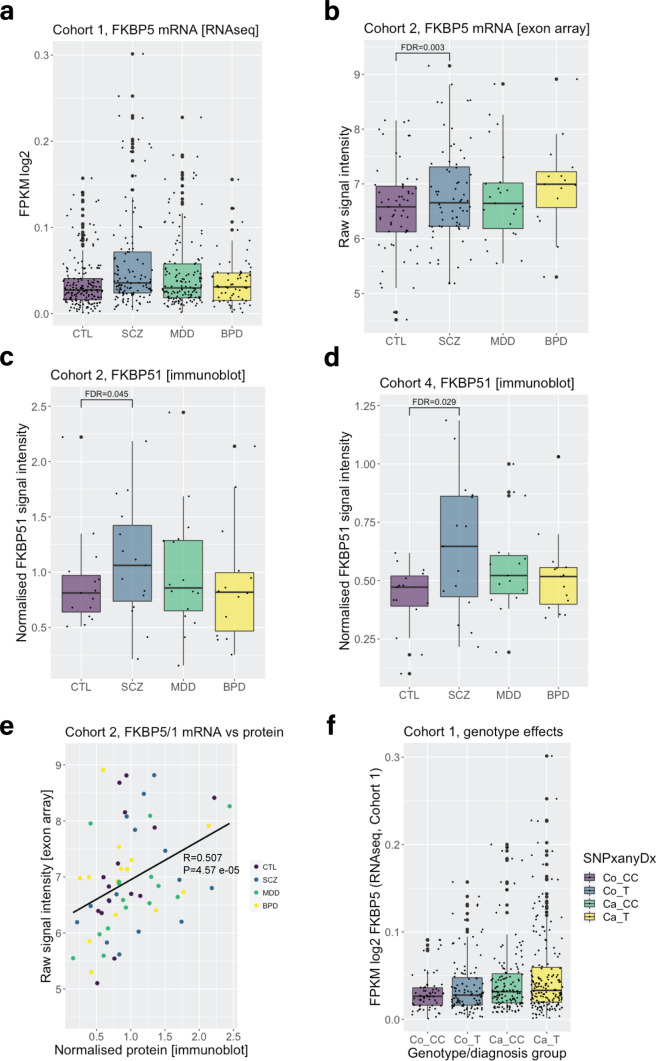
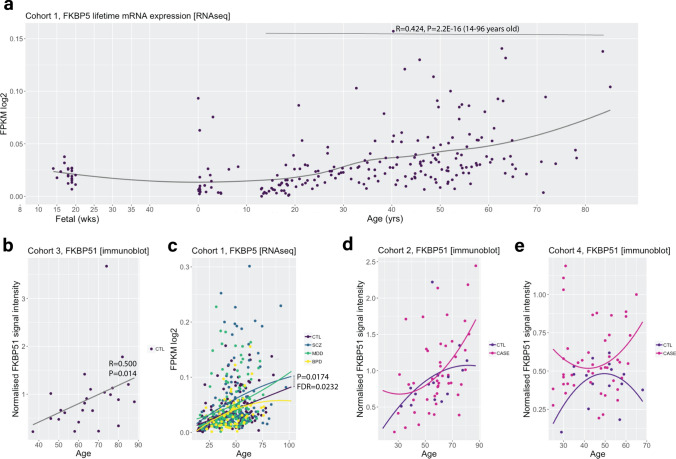
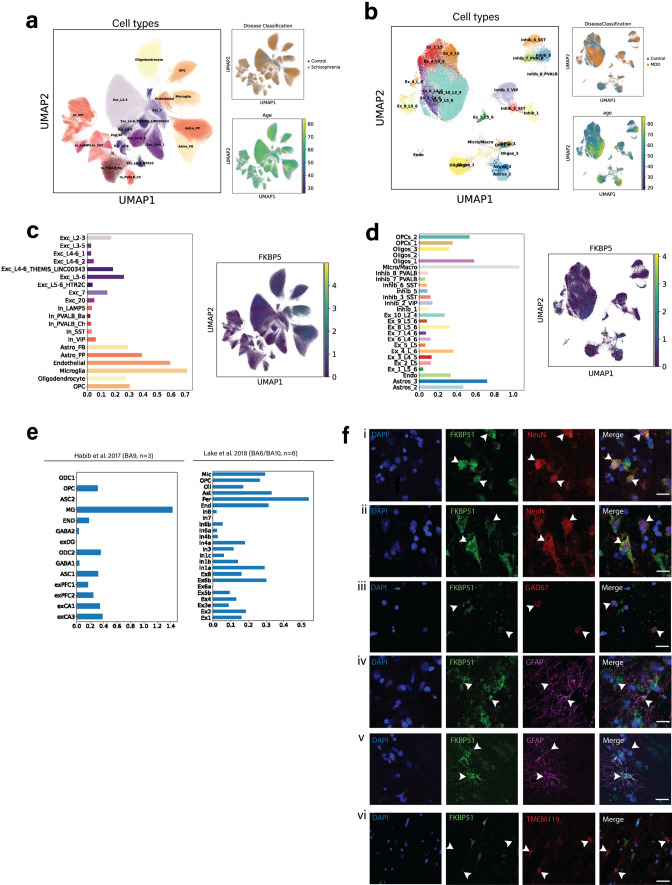
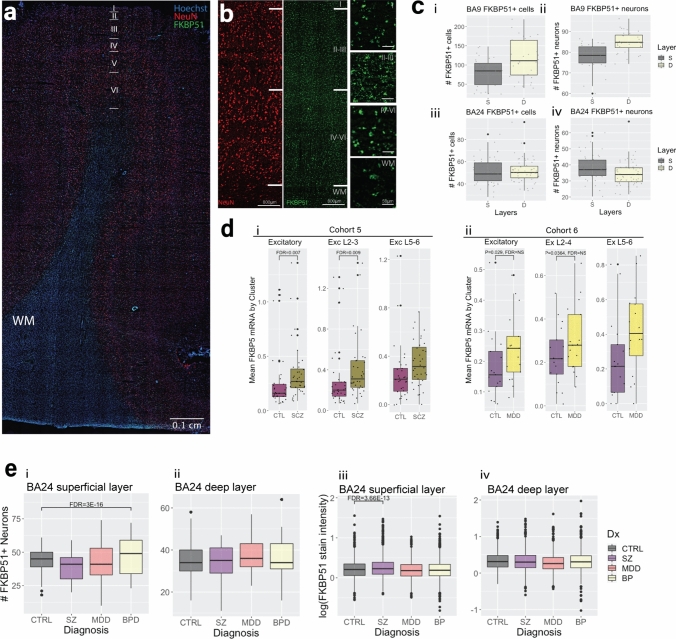
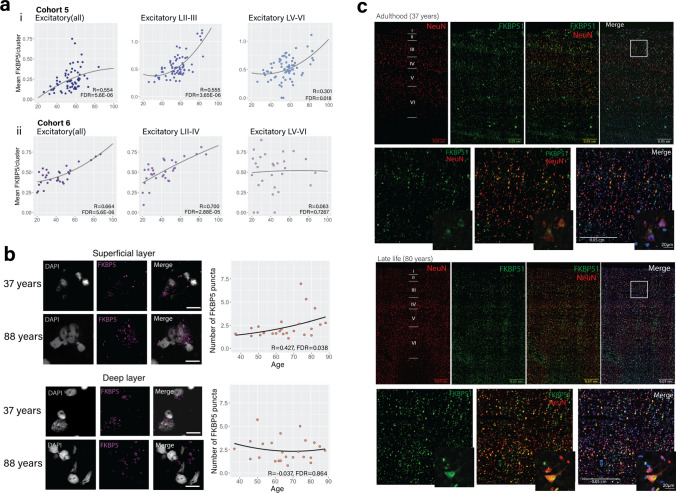
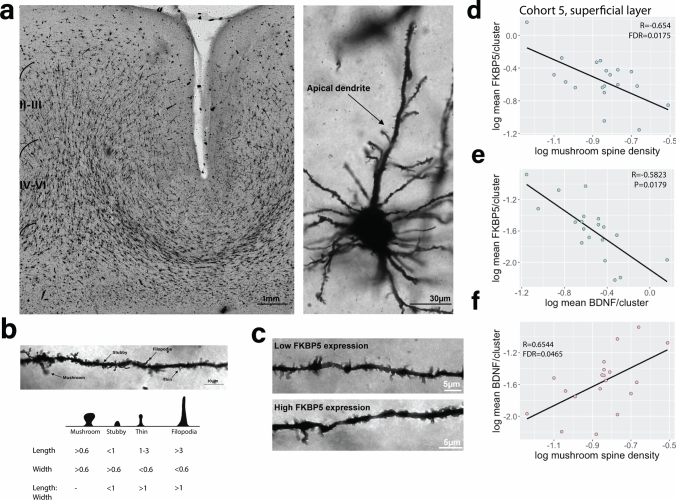
Identification and characterisation of novel targets for treatment is a priority in the field of psychiatry. FKBP5 is a gene with decades of evidence suggesting its pathogenic role in a subset of psychiatric patients, with potential to be leveraged as a therapeutic target for these individuals. While it is widely reported that FKBP5/FKBP51 mRNA/protein (FKBP5/1) expression is impacted by psychiatric disease state, risk genotype and age, it is not known in which cell types and sub-anatomical areas of the human brain this occurs. This knowledge is critical to propel FKBP5/1-targeted treatment development. Here, we performed an extensive, large-scale postmortem study (n = 1024) of FKBP5/1, examining neocortical areas (BA9, BA11 and ventral BA24/BA24a) derived from subjects that lived with schizophrenia, major depression or bipolar disorder. With an extensive battery of RNA (bulk RNA sequencing, single-nucleus RNA sequencing, microarray, qPCR, RNAscope) and protein (immunoblot, immunohistochemistry) analysis approaches, we thoroughly investigated the effects of disease state, ageing and genotype on cortical FKBP5/1 expression including in a cell type-specific manner. We identified consistently heightened FKBP5/1 levels in psychopathology and with age, but not genotype, with these effects strongest in schizophrenia. Using single-nucleus RNA sequencing (snRNAseq; BA9 and BA11) and targeted histology (BA9, BA24a), we established that these disease and ageing effects on FKBP5/1 expression were most pronounced in excitatory superficial layer neurons of the neocortex, and this effect appeared to be consistent in both the granular and agranular areas examined. We then found that this increase in FKBP5 levels may impact on synaptic plasticity, as FKBP5 gex levels strongly and inversely correlated with dendritic mushroom spine density and brain-derived neurotrophic factor (BDNF) levels in superficial layer neurons in BA11. These findings pinpoint a novel cellular and molecular mechanism that has potential to open a new avenue of FKBP51 drug development to treat cognitive symptoms in psychiatric disorders.
Keywords: Ageing; Depression; FKBP5; FKBP51; Postmortem brain; Psychosis; Single cell; Stress.
© 2023. The Author(s).
Conflict of interest statement
Dr Binder is a co-inventor of the following patent applications: FKBP5: a novel target for antidepressant therapy. European Patent # EP1687443 B1: Polymorphisms in ABCB1 associated with a lack of clinical response to medicaments. United States Patent # 8030033; Means and methods for diagnosing predisposition for treatment emergent suicidal ideation (TESI). European application number: 08016477.5, international application number: PCT/EP2009/061575. Dr Falkai has been an honorary speaker for AstraZeneca, Bristol Myers Squibb, Eli Lilly, Essex, GE Healthcare, GlaxoSmithKline, Janssen Cilag, Lundbeck, Otsuka, Pfizer, Servier and Takeda. During the past 5 years, but not presently, Dr Falkai has been a member of the advisory boards of Janssen-Cilag, AstraZeneca, Eli Lilly and Lundbeck. Dr Schmitt has been an honorary speaker for TAD Pharma and Roche and has been a member of advisory boards for Roche. The remaining authors declare no competing interests or conflicts of interest.
Figures
Comment in
-
Molecular integrators of stress and aging: the example of FKBP5.Acta Neuropathol. 2023 Jun;145(6):713-715. doi: 10.1007/s00401-023-02572-2. Epub 2023 Apr 5. Acta Neuropathol. 2023. PMID: 37020089 No abstract available.
References
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc Ser B (Methodol) 1995;57:289–300.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
