

Regulation of mTOR by phosphatidic acid
- PMID: 36732094
- PMCID: PMC9957947
- DOI: 10.1016/j.tem.2023.01.004
Regulation of mTOR by phosphatidic acid
Abstract
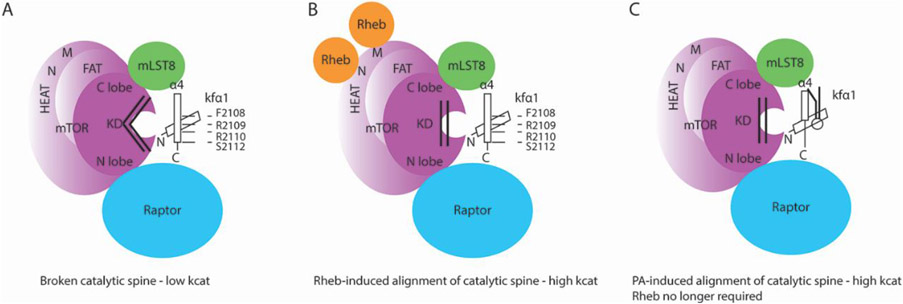
mTORC1, the mammalian target of rapamycin complex 1, is a key regulator of cellular physiology. The lipid metabolite phosphatidic acid (PA) binds to and activates mTORC1 in response to nutrients and growth factors. We review structural findings and propose a model for PA activation of mTORC1. PA binds to a highly conserved sequence in the α4 helix of the FK506 binding protein 12 (FKBP12)/rapamycin-binding (FRB) domain of mTOR. It is proposed that PA binding to two adjacent positively charged amino acids breaks and shortens the C-terminal region of helix α4. This has profound consequences for both substrate binding and the catalytic activity of mTORC1.
Keywords: FRB; Rheb; mTOR; phosphatidic acid; phospholipase D; structure.
Copyright © 2023 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
