

Kinetics of cone specific G-protein signaling in avian photoreceptor cells
- PMID: 36733826
- PMCID: PMC9887155
- DOI: 10.3389/fnmol.2023.1107025
Kinetics of cone specific G-protein signaling in avian photoreceptor cells
Abstract
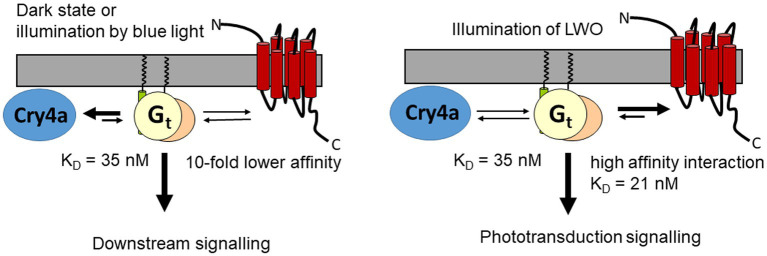
Cone photoreceptor cells of night-migratory songbirds seem to process the primary steps of two different senses, vision and magnetoreception. The molecular basis of phototransduction is a prototypical G protein-coupled receptor pathway starting with the photoexcitation of rhodopsin or cone opsin thereby activating a heterotrimeric G protein named transducin. This interaction is well understood in vertebrate rod cells, but parameter describing protein-protein interactions of cone specific proteins are rare and not available for migratory birds. European robin is a model organism for studying the orientation of birds in the earth magnetic field. Recent findings showed a link between the putative magnetoreceptor cryptochrome 4a and the cone specific G-protein of European robin. In the present work, we investigated the interaction of European robin cone specific G protein and cytoplasmic regions of long wavelength opsin. We identified the second loop in opsin connecting transmembrane regions three and four as a critical binding interface. Surface plasmon resonance studies using a synthetic peptide representing the second cytoplasmic loop and purified G protein α-subunit showed a high affinity interaction with a K D value of 21 nM. Truncation of the G protein α-subunit at the C-terminus by six amino acids slightly decreased the affinity. Our results suggest that binding of the G protein to cryptochrome can compete with the interaction of G protein and non-photoexcited long wavelength opsin. Thus, the parallel presence of two different sensory pathways in bird cone photoreceptors is reasonable under dark-adapted conditions or during illumination with short wavelengths.
Keywords: G protein; cone opsin; cone outer segment; photoreceptor; protein–protein interaction.
Copyright © 2023 Yee, Görtemaker, Wellpott and Koch.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
-
- Alves I. D., Salgado G. F., Salamon Z., Brown M. F., Tollin G., Hruby V. J. (2005). Phosphatidylethanolamine enhances rhodopsin photoactivation and transducin binding in a solid supported lipid bilayer as determined using plasmon-waveguide resonance spectroscopy. Biophys. J. 88, 198–210. doi: 10.1529/biophysj.104.046722, PMID: - DOI - PMC - PubMed
-
- Bolte P., Einwich A., Seth P. K., Chetverikova R., Heyers D., Wojahn I., et al. (2021). Cryptochrome 1a localisation in light-and dark-adapted retinae of several migratory and non-migratory bird species: no signs of light-dependent activation. Ethol. Ecol. Evol. 33, 248–272. doi: 10.1080/03949370.2020.1870571 - DOI
LinkOut - more resources
Full Text Sources