

The dengue virus 4 component of NIAID's tetravalent TV003 vaccine drives its innate immune signature
- PMID: 36734144
- PMCID: PMC9899989
- DOI: 10.1177/15353702231151241
The dengue virus 4 component of NIAID's tetravalent TV003 vaccine drives its innate immune signature
Abstract
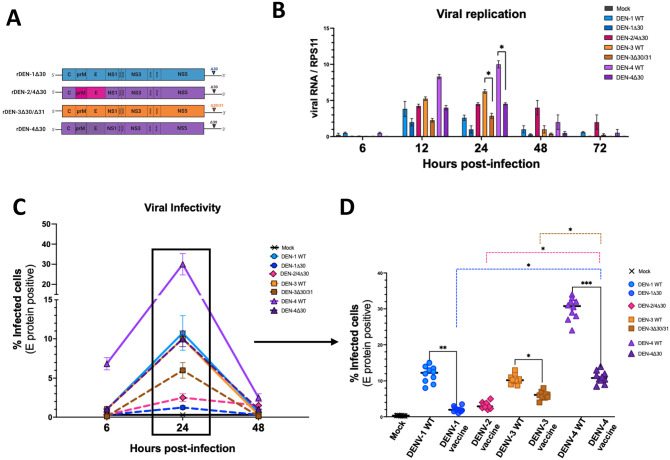
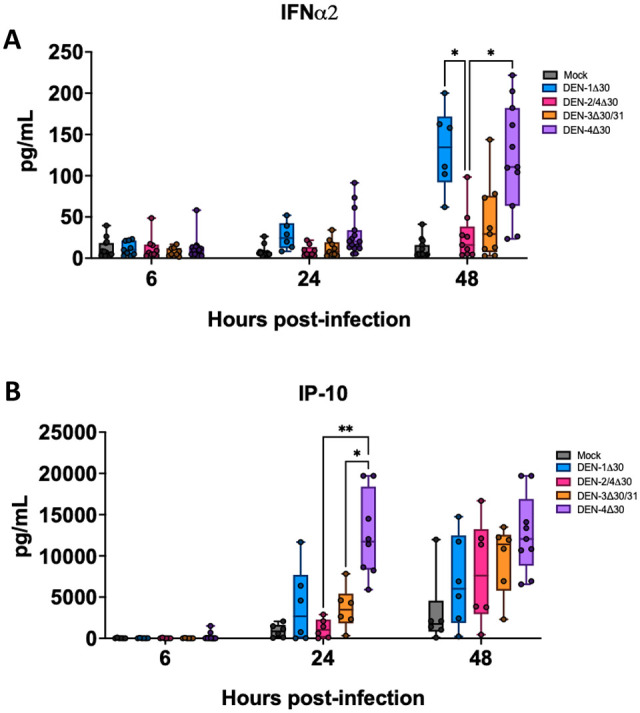
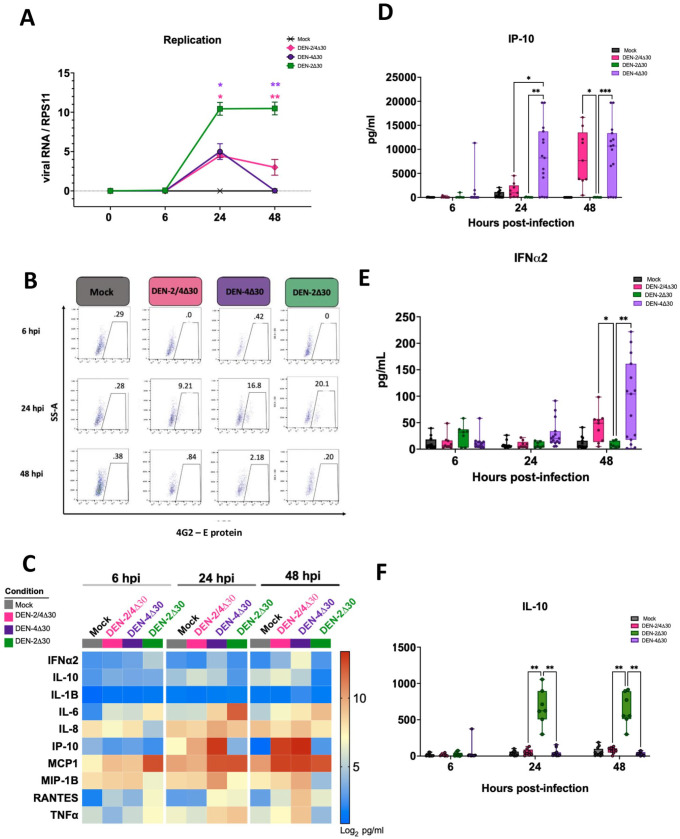
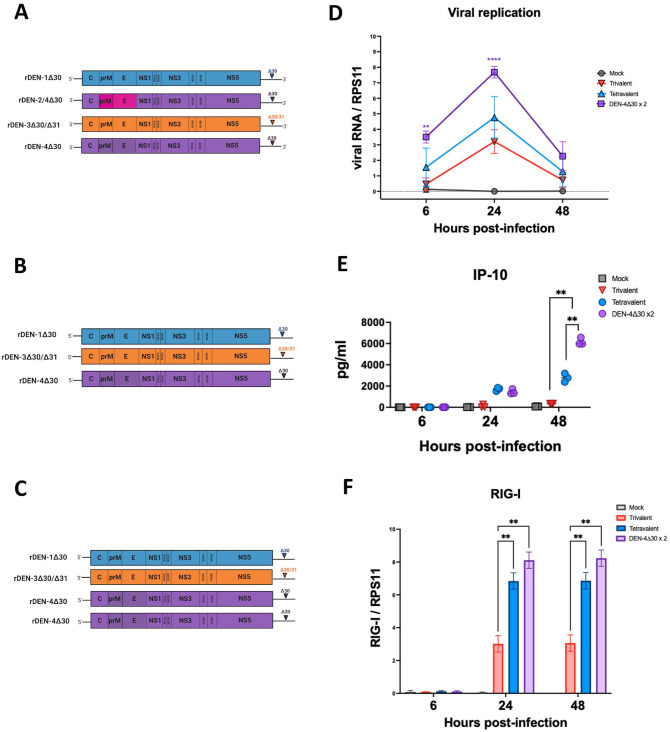
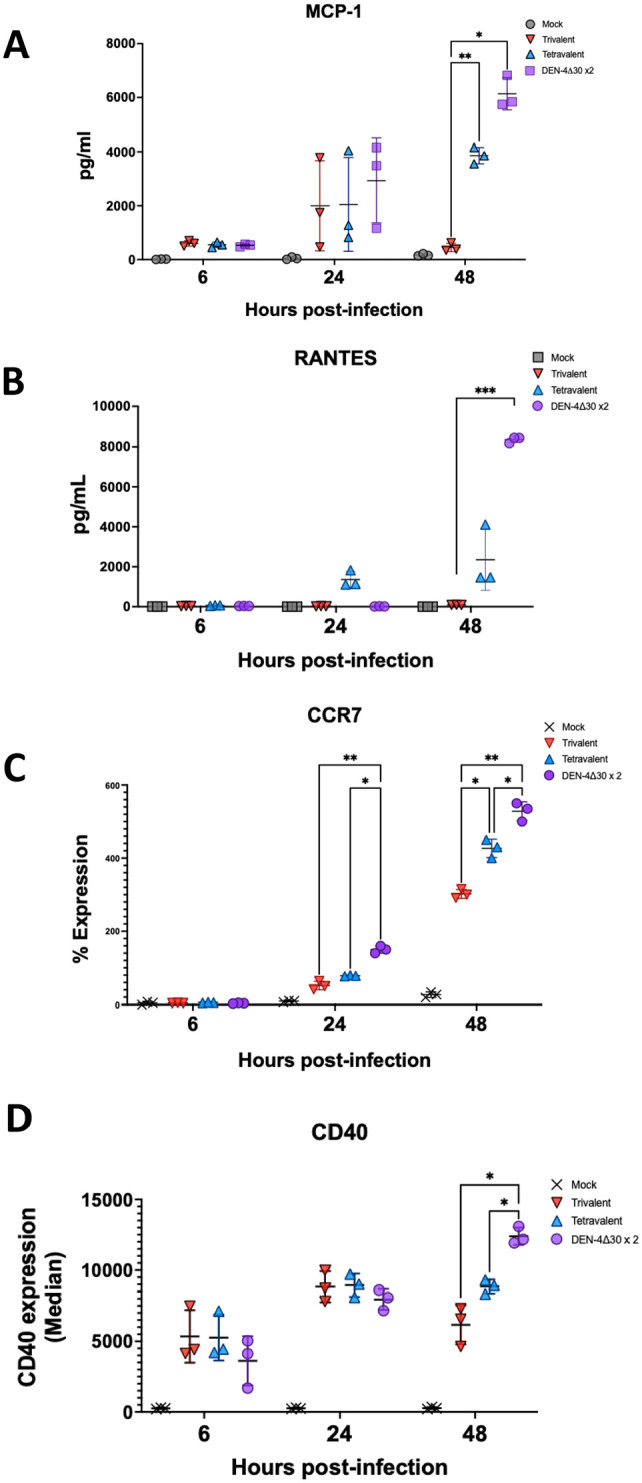
Annually, roughly 2.5 billion people are at risk for dengue virus (DENV) infection, and the incidence of infection has increased 30-fold since its discovery in the 1900s. At present, there are no globally licensed antiviral treatments or vaccines that protect against all four of the DENV serotypes. The NIAID Live Attenuated Tetravalent Vaccine (LATV) dengue vaccine candidate is composed of variants of three DENV serotypes attenuated by a 30 nucleotide (Δ30) deletion in the 3' untranslated region and a fourth component that is a chimeric virus in which the prM and E genes of DENV-2 replace those of DENV-4 on the rDEN4Δ30 backbone. The vaccine candidate encodes the non-structural proteins of DENV-1, DENV-3, and DENV-4, which could be of critical importance in the presentation of DENV-specific epitopes in a manner that facilitates antigen presentation and confers higher protection. Our findings demonstrate that the attenuation mechanism (Δ30) resulted in decreased viral infectivity and replication for each vaccine virus in monocyte-derived dendritic cells but were able to generate a robust innate immune response. When tested as monovalent viruses, DEN-4Δ30 displayed the most immunogenic profile. In addition, we found that the tetravalent DENV formulation induced a significantly greater innate immune response than the trivalent formulation. We demonstrate that the presence of two components with a DENV-4Δ30 backbone is necessary for the induction of RANTES, CD40, IP-10, and Type I IFN by the tetravalent formulation. Finally, we found that the DEN-4Δ30 backbone in the DENV-2 component of the vaccine enhanced its antigenic properties, as evidenced by enhanced ability to induce IP-10 and IFNα2 in monocyte-derived dendritic cells. In sum, our study shows that the Δ30 and Δ30/Δ31 mutations attenuate the DENV vaccine strains in terms of replication and infectivity while still allowing the induction of a robust innate immune response.
Keywords: Dengue; innate immunity; vaccine; vaccine efficacy.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
References
-
- Weiskopf D, Angelo MA, Bangs DJ, Sidney J, Paul S, Peters B, de Silva AD, Lindow JC, Diehl SA, Whitehead S, Durbin A, Kirkpatrick B, Sette A. The human CD8+ T cell responses induced by a live attenuated tetravalent dengue vaccine are directed against highly conserved epitopes. J Virol 2015;89(1):120–8 - PMC - PubMed
-
- Centers for Disease Control and Prevention. Areas with risk of dengue, 2021, https://www.cdc.gov/dengue/areaswithrisk/index.html
-
- WHO. Dengue and severe dengue, 2022, https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials