

Lineage Tracing and Single-Nucleus Multiomics Reveal Novel Features of Adaptive and Maladaptive Repair after Acute Kidney Injury
- PMID: 36735940
- PMCID: PMC10103206
- DOI: 10.1681/ASN.0000000000000057
Lineage Tracing and Single-Nucleus Multiomics Reveal Novel Features of Adaptive and Maladaptive Repair after Acute Kidney Injury
Abstract
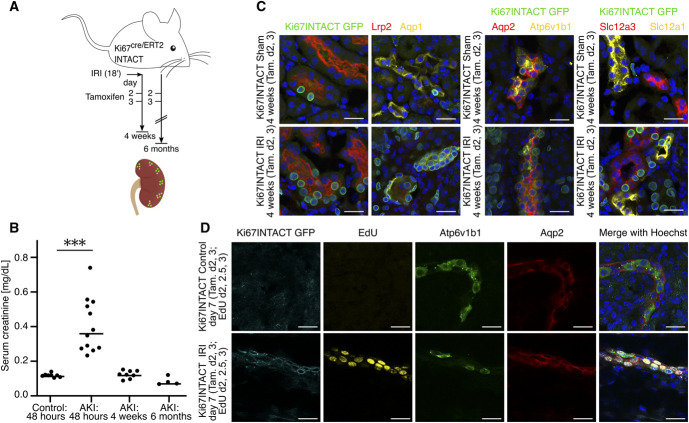
Significance statement: Understanding the mechanisms underlying adaptive and maladaptive renal repair after AKI and their long-term consequences is critical to kidney health. The authors used lineage tracing of cycling cells and single-nucleus multiomics (profiling transcriptome and chromatin accessibility) after AKI. They demonstrated that AKI triggers a cell-cycle response in most epithelial and nonepithelial kidney cell types. They also showed that maladaptive proinflammatory proximal tubule cells (PTCs) persist until 6 months post-AKI, although they decreased in abundance over time, in part, through cell death. Single-nucleus multiomics of lineage-traced cells revealed regulatory features of adaptive and maladaptive repair. These included activation of cell state-specific transcription factors and cis-regulatory elements, and effects in PTCs even after adaptive repair, weeks after the injury event.
Background: AKI triggers a proliferative response as part of an intrinsic cellular repair program, which can lead to adaptive renal repair, restoring kidney structure and function, or maladaptive repair with the persistence of injured proximal tubule cells (PTCs) and an altered kidney structure. However, the cellular and molecular understanding of these repair programs is limited.
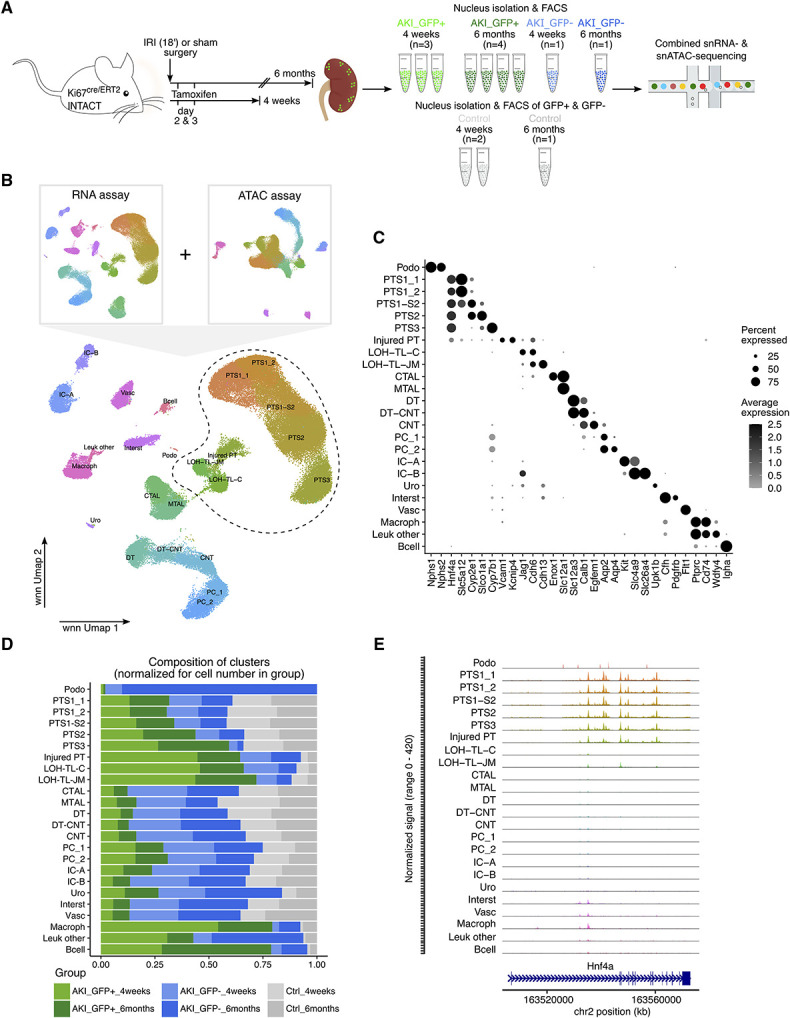
Methods: To examine chromatin and transcriptional responses in the same cell upon ischemia-reperfusion injury (IRI), we combined genetic fate mapping of cycling ( Ki67+ ) cells labeled early after IRI with single-nucleus multiomics-profiling transcriptome and chromatin accessibility in the same nucleus-and generated a dataset of 83,315 nuclei.
Results: AKI triggered a broad cell cycle response preceded by cell type-specific and global transcriptional changes in the nephron, the collecting and vascular systems, and stromal and immune cell types. We observed a heterogeneous population of maladaptive PTCs throughout proximal tubule segments 6 months post-AKI, with a marked loss of maladaptive cells from 4 weeks to 6 months. Gene expression and chromatin accessibility profiling in the same nuclei highlighted differences between adaptive and maladaptive PTCs in the activity of cis-regulatory elements and transcription factors, accompanied by corresponding changes in target gene expression. Adaptive repair was associated with reduced expression of genes encoding transmembrane transport proteins essential to kidney function.
Conclusions: Analysis of genome organization and gene activity with single-cell resolution using lineage tracing and single-nucleus multiomics offers new insight into the regulation of renal injury repair. Weeks to months after mild-to-moderate IRI, maladaptive PTCs persist with an aberrant epigenetic landscape, and PTCs exhibit an altered transcriptional profile even following adaptive repair.
Copyright © 2023 by the American Society of Nephrology.
Conflict of interest statement
B.D.H. is a consultant Chinook Therapeutics, Janssen Research & Development, LLC, and Pfizer; holds equity in Chinook Therapeutics; and has had grant support from Chinook Therapeutics, Janssen and Pfizer (within last 3 years). B. Humphreys also reports Patents or Royalties: Evotec, AG; Advisory or Leadership Role: American Journal of Physiology Renal Physiology Editorial Board, ASCI President-elect, Board of Scientific Advisors, Chinook Therapeutics SAB, JCI Insight Editorial Board, Kidney International Editorial Board, Regenerative Medicine Crossing Borders SAB, RegMed XB, and Seminars in Nephrology Editorial Board.
A.P.M. receives consulting fees or stock options for his scientific advisory role for eGENESIS, IVIVA Medical, Novartis, and TRESTLE BioTherapeutics. A. McMahon also reports research funding: Amgen; Patents or Royalties: Hedgehog technology licenses through Harvard to Curis, received license/royalty payments from Curis; Advisory or Leadership Role: California Institute of Technology—Jacobs Institute, Max Planck Institute Molecular Genetics Berlin; and Other Interests or Relationships: Committee of University Kidney Research Organization (UKRO). J. van Gestel reports Employer: EMBL. All remaining authors have nothing to disclose.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
