

Impaired Endogenous Neurosteroid Signaling Contributes to Behavioral Deficits Associated With Chronic Stress
- PMID: 36736870
- PMCID: PMC10363189
- DOI: 10.1016/j.biopsych.2023.01.022
Impaired Endogenous Neurosteroid Signaling Contributes to Behavioral Deficits Associated With Chronic Stress
Abstract
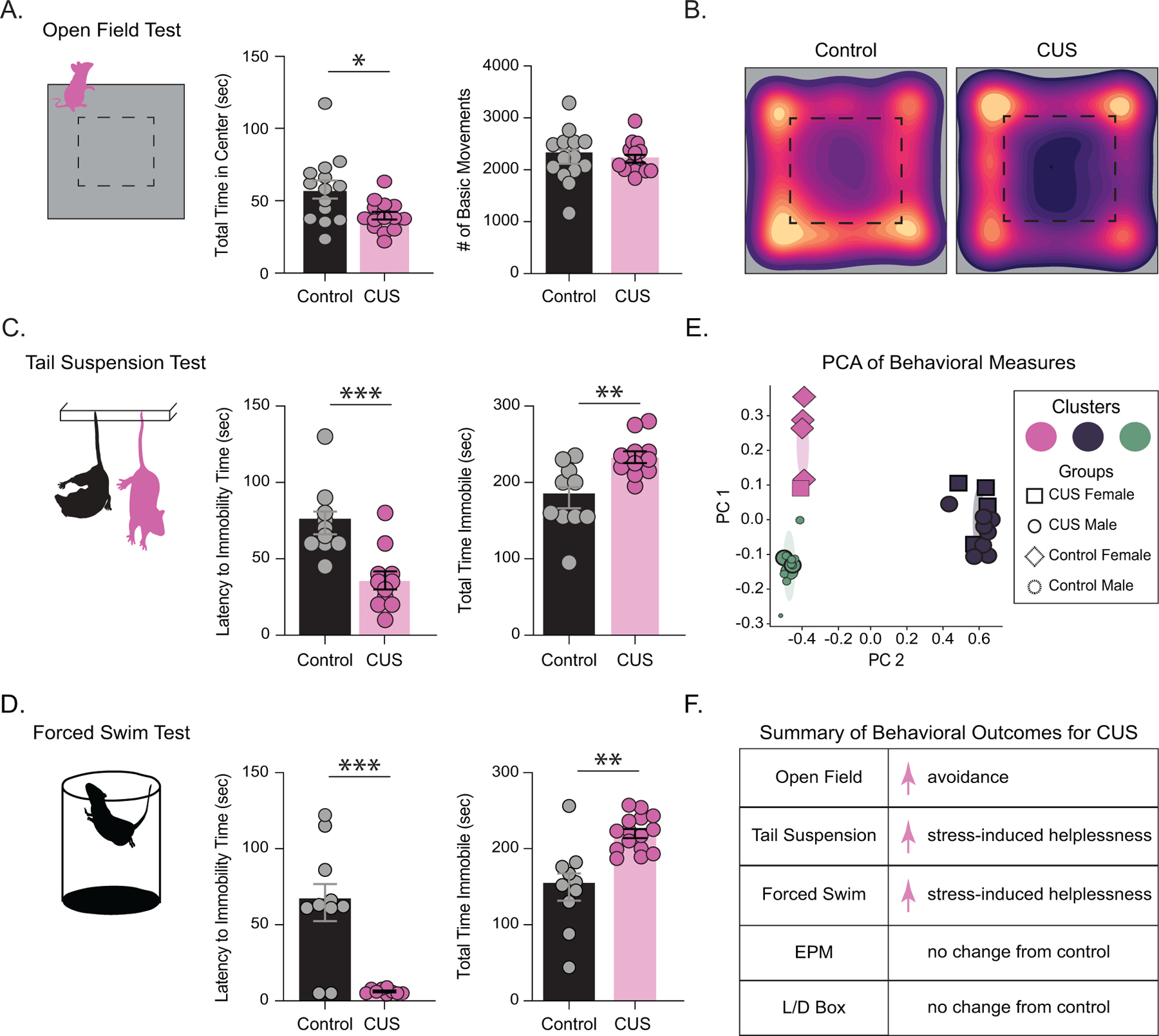
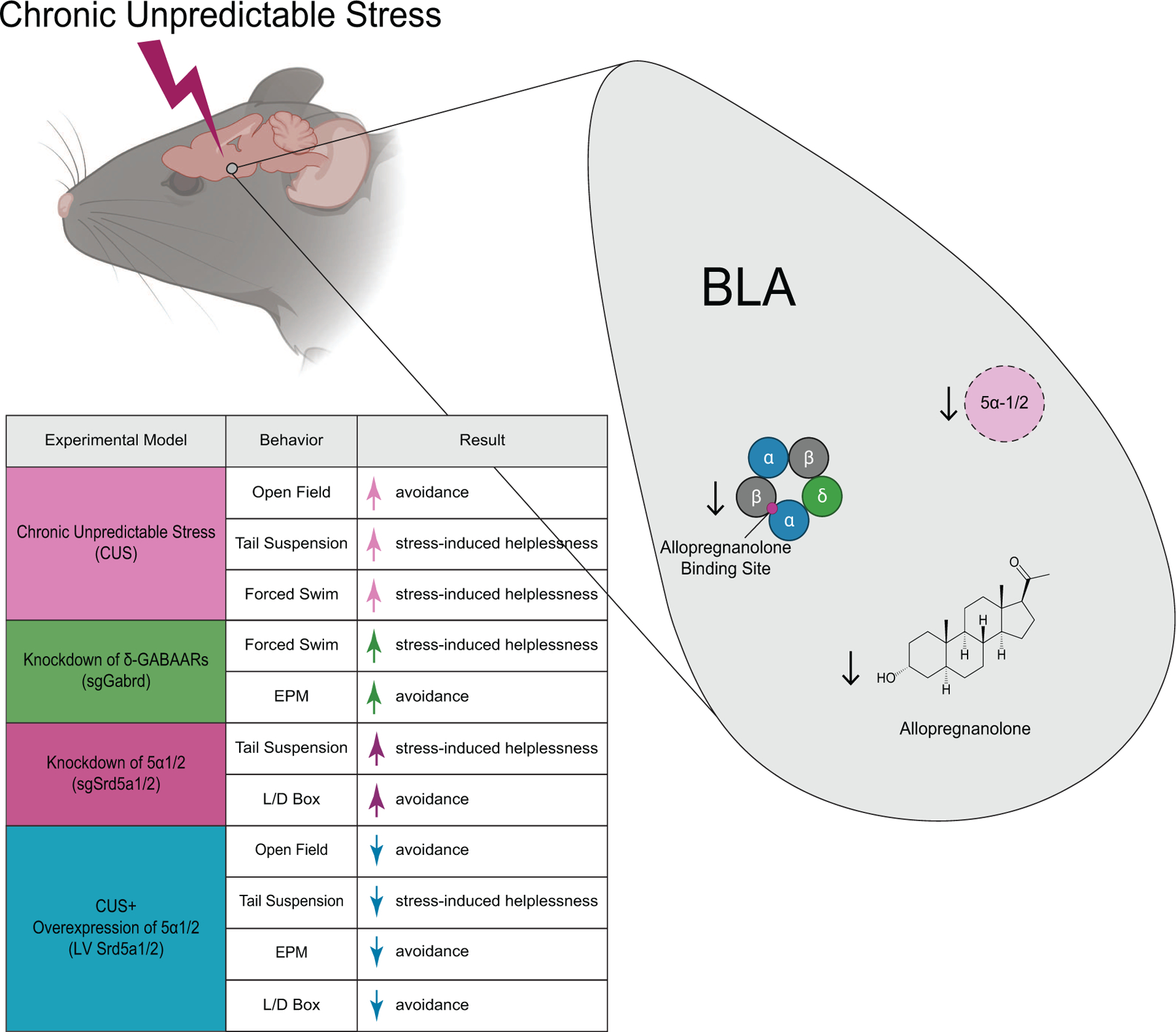
Background: Chronic stress is a major risk factor for psychiatric illnesses, including depression. However, the pathophysiological mechanisms whereby stress leads to mood disorders remain unclear. Allopregnanolone acts as a positive allosteric modulator preferentially on δ subunit-containing GABAA (gamma-aminobutyric acid A) receptors. Accumulating clinical and preclinical evidence supports the antidepressant effects of exogenous administration of allopregnanolone analogs; yet, the role of endogenous allopregnanolone in the pathophysiology of depression remains unknown.
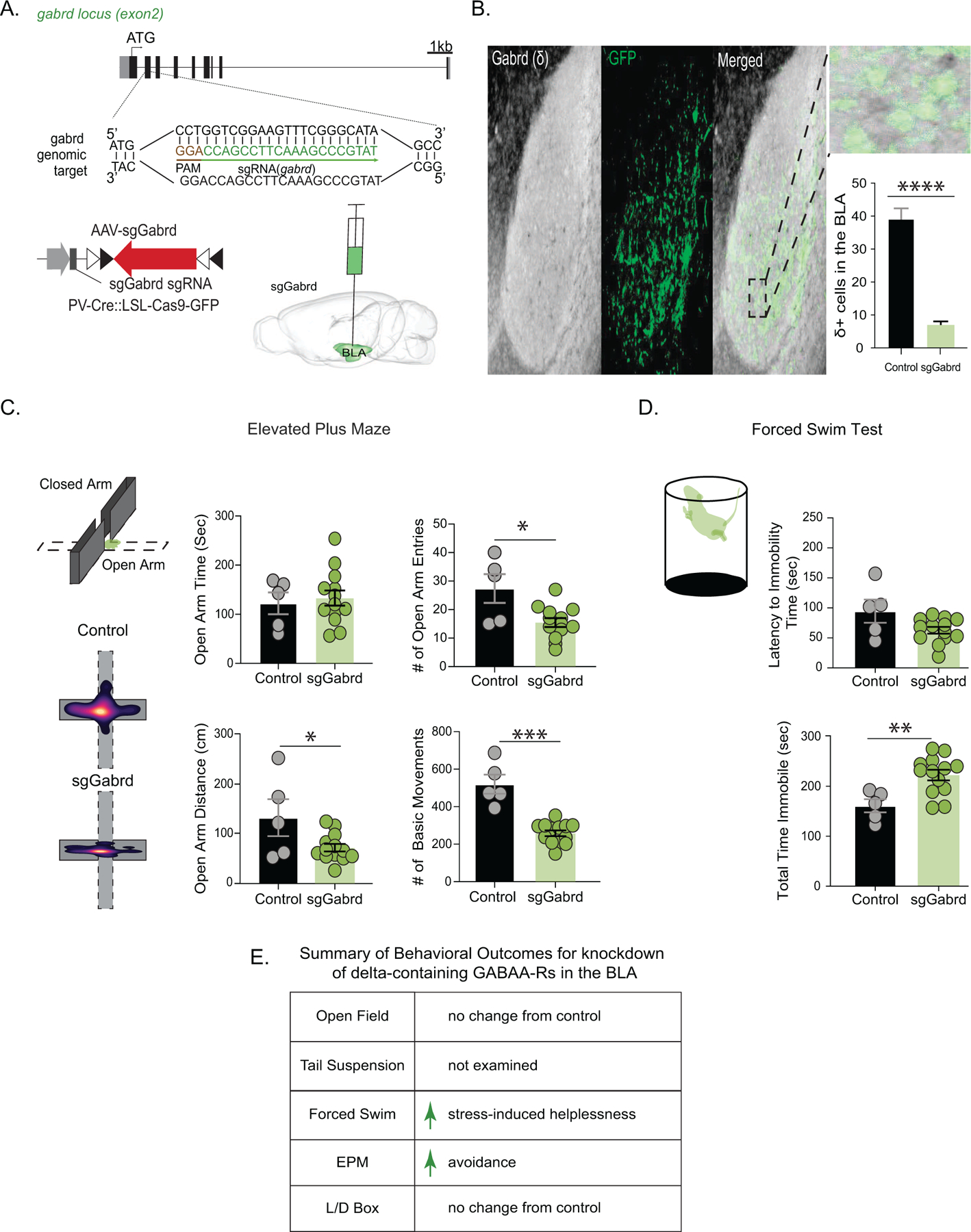
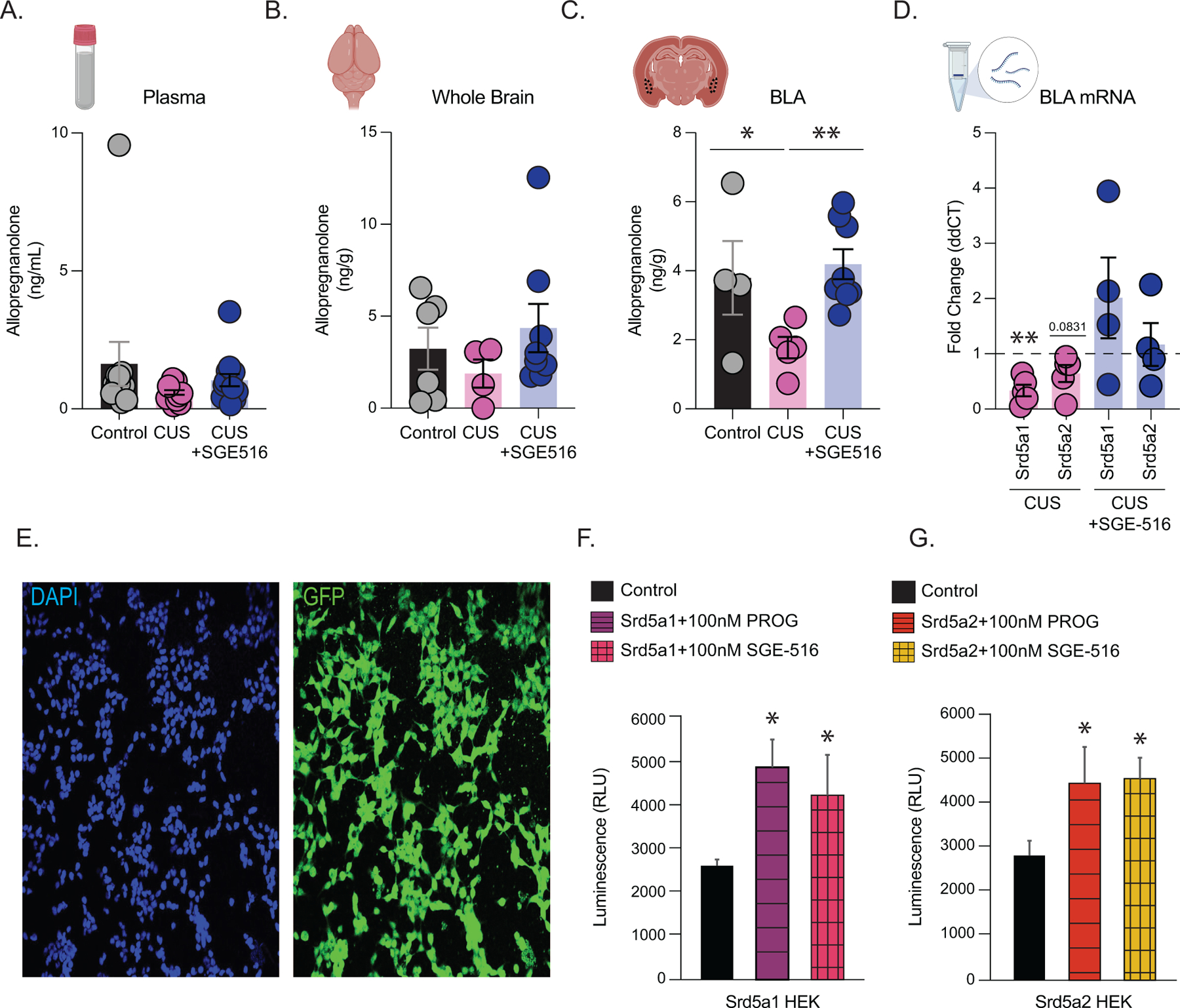
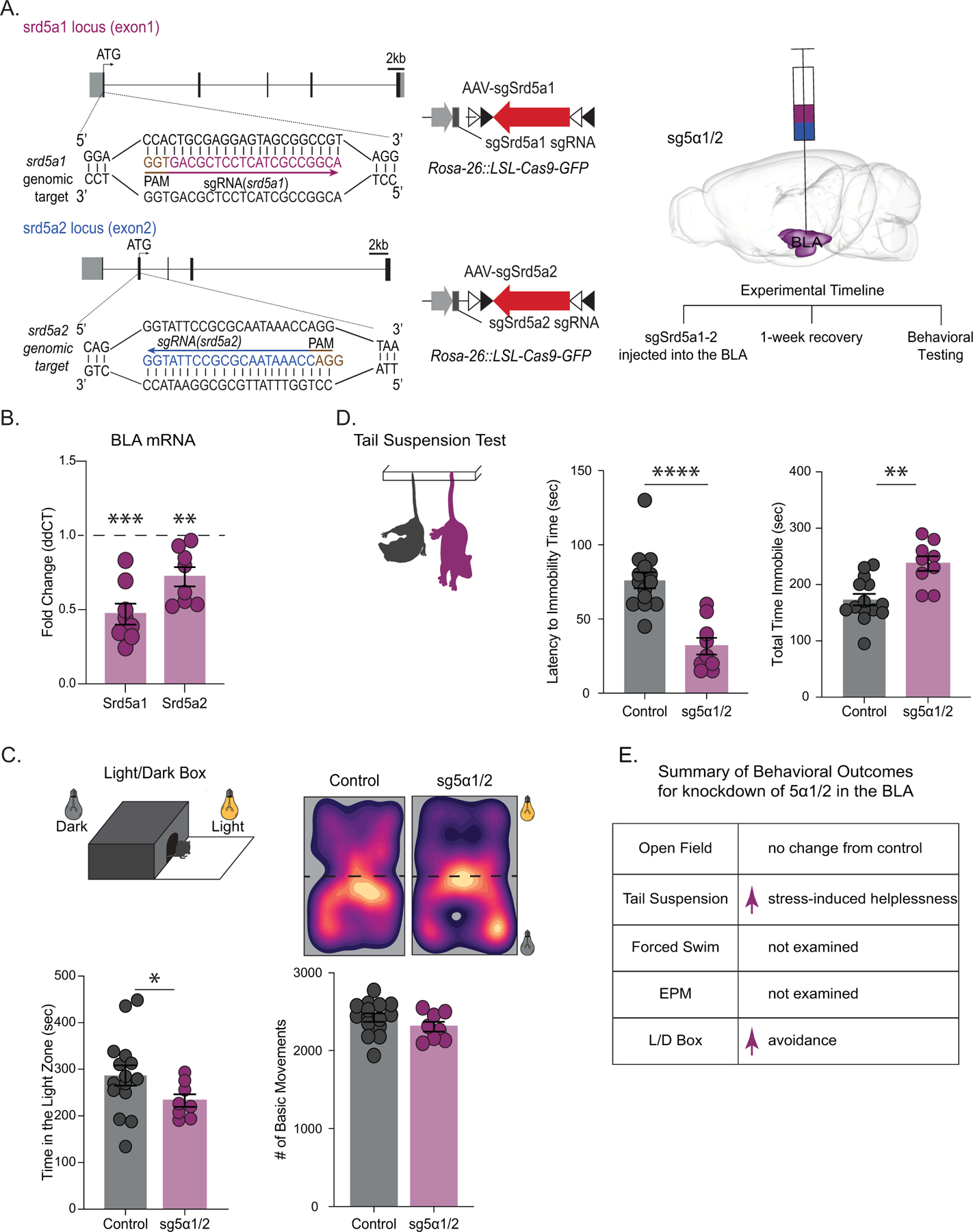
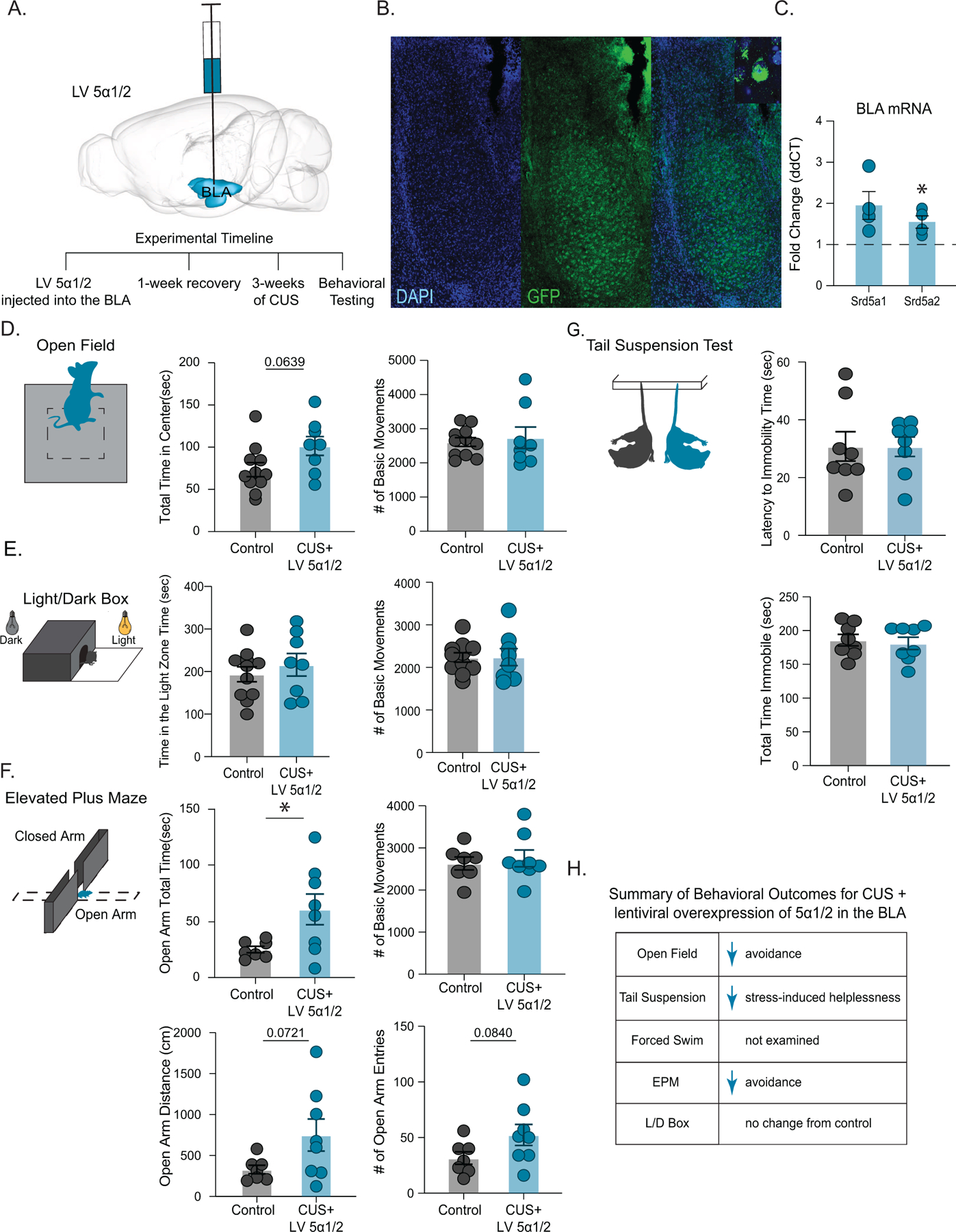
Methods: We utilized a chronic unpredictable stress (CUS) mouse model, followed by behavioral and biochemical assays, to examine whether altered neurosteroid signaling contributes to behavioral outcomes following CUS. We subsequently performed in vivo CRISPR (clustered regularly interspaced short palindromic repeats) knockdown of rate-limiting enzymes involved in allopregnanolone synthesis, 5α-reductase type 1 and 2 (5α1/2), in addition to lentiviral overexpression of 5α1/2 in the basolateral amygdala (BLA) of mice that underwent CUS to assess the impact of 5α1/2 on behavioral outcomes.
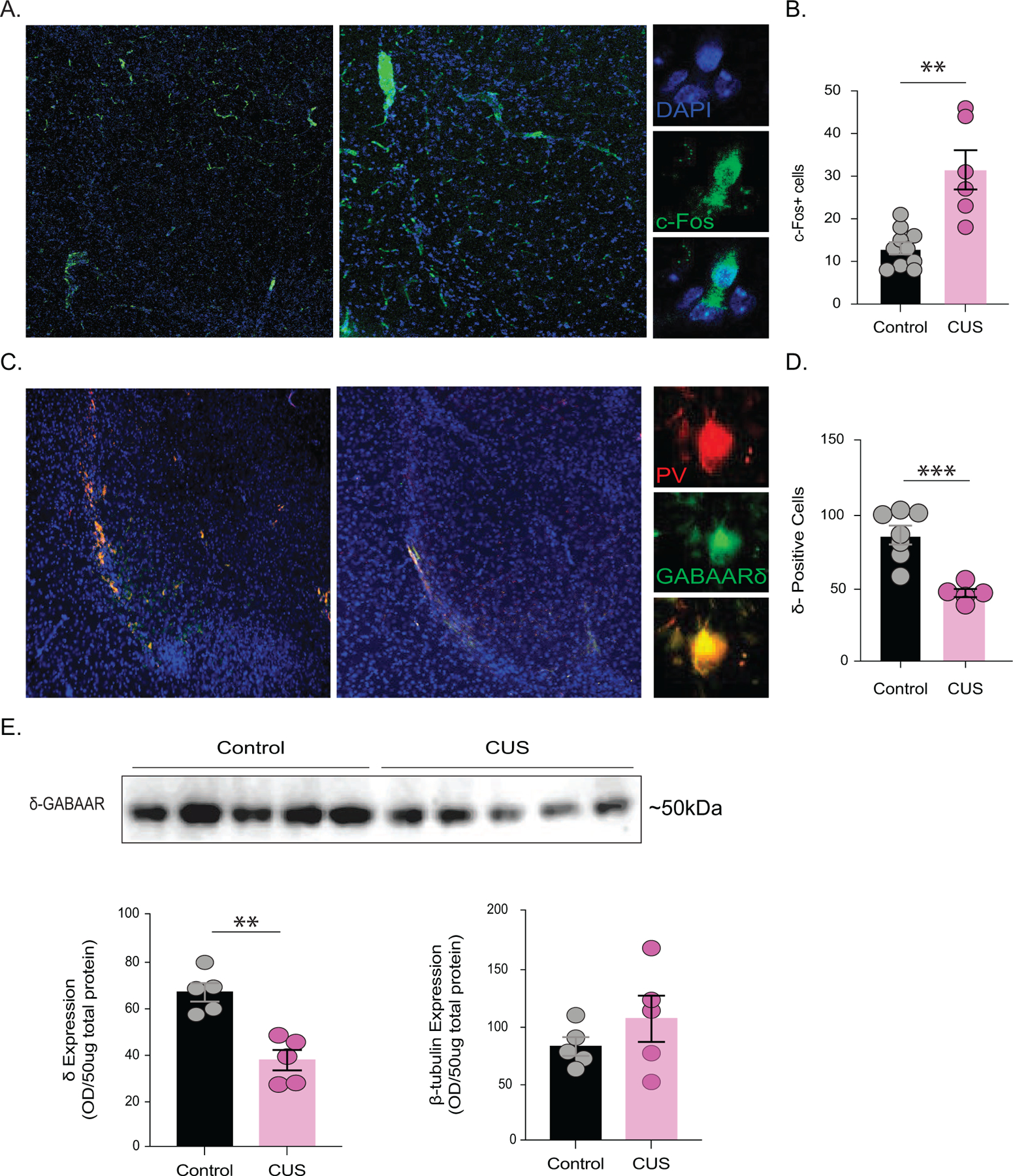
Results: The expression of δ subunit-containing GABAA receptors and endogenous levels of allopregnanolone were reduced in the BLA following CUS. Treatment with an exogenous allopregnanolone analog, SGE-516, was sufficient to increase allopregnanolone levels in the BLA following CUS. Knockdown of 5α1/2 in the BLA mimicked the behavioral outcomes associated with CUS. Conversely, overexpression of 5α1/2 in the BLA improved behavioral outcomes following CUS.
Conclusions: Our findings demonstrate that chronic stress impairs endogenous neurosteroid signaling in the BLA, which is sufficient to induce behavioral deficits. Further, these studies suggest that allopregnanolone-based treatments may directly target the underlying pathophysiology of mood disorders suggesting that targeting endogenous neurosteroidogenesis may offer a novel therapeutic strategy.
Keywords: Allopregnanolone; Depression; GABA; Neurosteroidogenesis; Neurosteroids; Stress.
Copyright © 2023 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Optimizing Neuroactive Steroid Treatments for Stress-Related Affective Symptoms: Brief Reflections on Patient Population, Timing, and Access.Biol Psychiatry. 2023 Aug 1;94(3):191-193. doi: 10.1016/j.biopsych.2023.05.015. Biol Psychiatry. 2023. PMID: 37437990 Free PMC article. No abstract available.
References
-
- Dong E, Matsumoto K, Uzunova V, Sugaya I, Takahata H, Nomura H, et al. (2001). Brain 5α-dihydroprogesterone and allopregnanolone synthesis in a mouse model of protracted social isolation. Proceedings of the National Academy of Sciences of the United States of America, 98(5), 2849–2854. 10.1073/pnas.051628598 - DOI - PMC - PubMed
-
- Serra M, Pisu MG, Littera M, Papi G, Sanna E, Tuveri F, et al. (2002). Social isolation-induced decreases in both the abundance of neuroactive steroids and Gabaa receptor function in rat brain. Journal of Neurochemistry, 75(2), 732–740. - PubMed
-
- Agís-Balboa RC, Pinna G, Pibiri F, Kadriu B, Costa E, & Guidotti A (2007). Down-regulation of neurosteroid biosynthesis in corticolimbic circuits mediates social isolation-induced behavior in mice. Proceedings of the National Academy of Sciences of the United States of America, 104(47), 18736–18741. 10.1073/pnas.0709419104 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
