

Intrinsic STING Switches off Pathogenetic Programs of Th1 Cells to Inhibit Colitis
- PMID: 36736893
- PMCID: PMC10040963
- DOI: 10.1016/j.jcmgh.2023.01.010
Intrinsic STING Switches off Pathogenetic Programs of Th1 Cells to Inhibit Colitis
Abstract
Background & aims: T helper 1 (Th1) effector cells are implicated in inflammatory bowel disease. The stimulator of interferon genes (STING), an intracellular DNA sensor, has been shown to regulate infection and various cancers. However, whether and how intrinsic STING signaling in Th1 cells regulates colitis is still unknown.
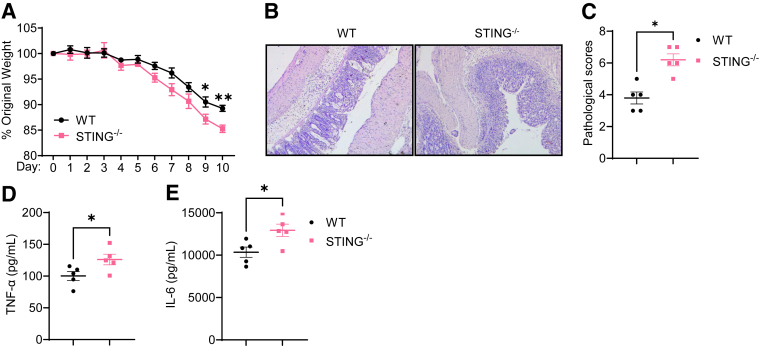
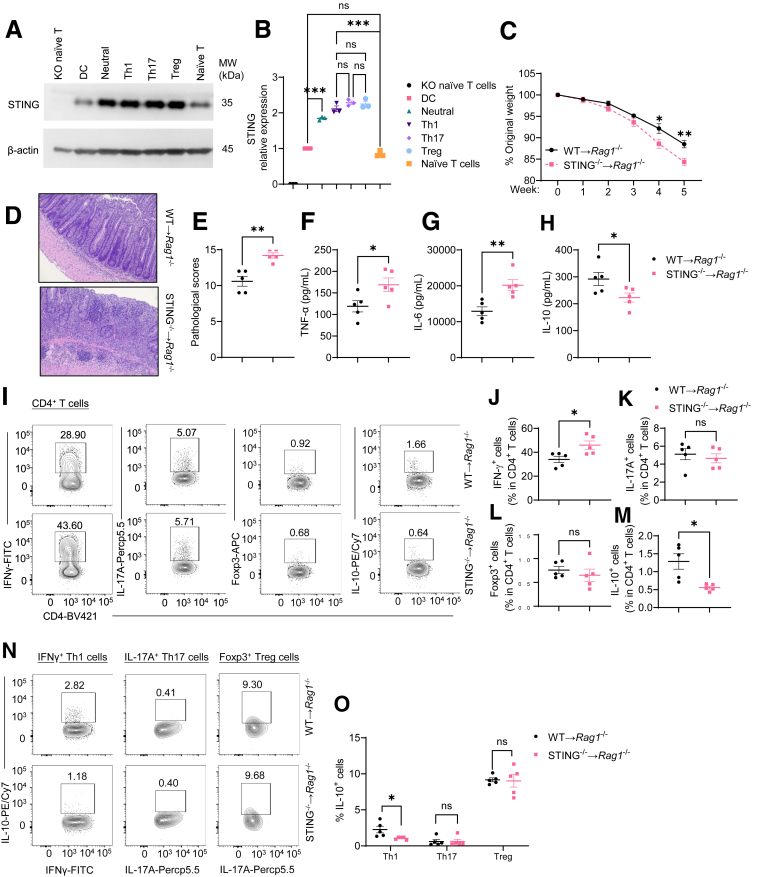
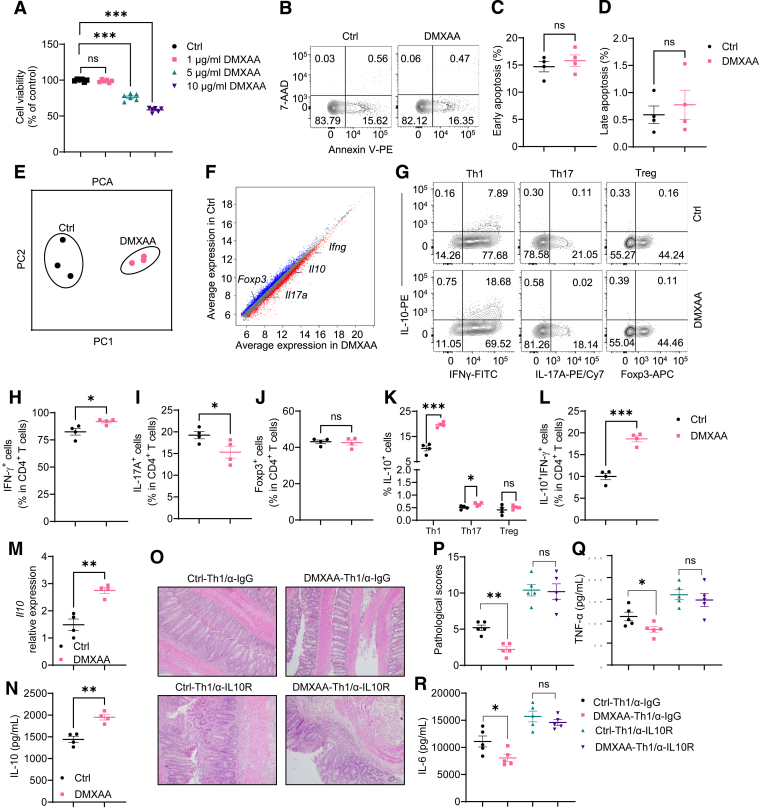
Methods: Dextran sodium sulfate-induced colitis and wild-type/STING-deficient CD4+T cell adoptive transfer models were used to analyze the role of STING in regulating colitis. The effect of STING on Th1 cells was determined by flow cytometry, RNA sequencing, metabolic assays, and mitochondrial functions. 16S ribosomal RNA sequencing and germ-free mice were used to investigate whether the microbiota were involved. The in vivo effect of STING agonist in murine colitis was determined. The expression and role of STING in human T cells were also determined.
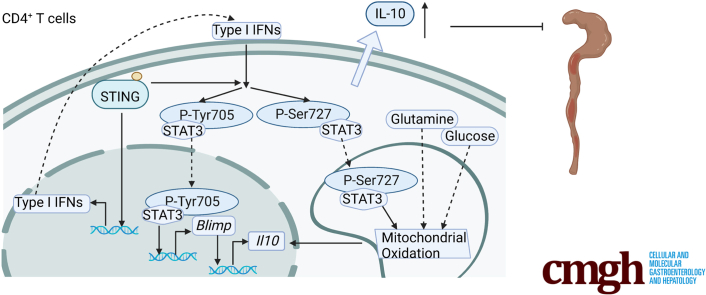
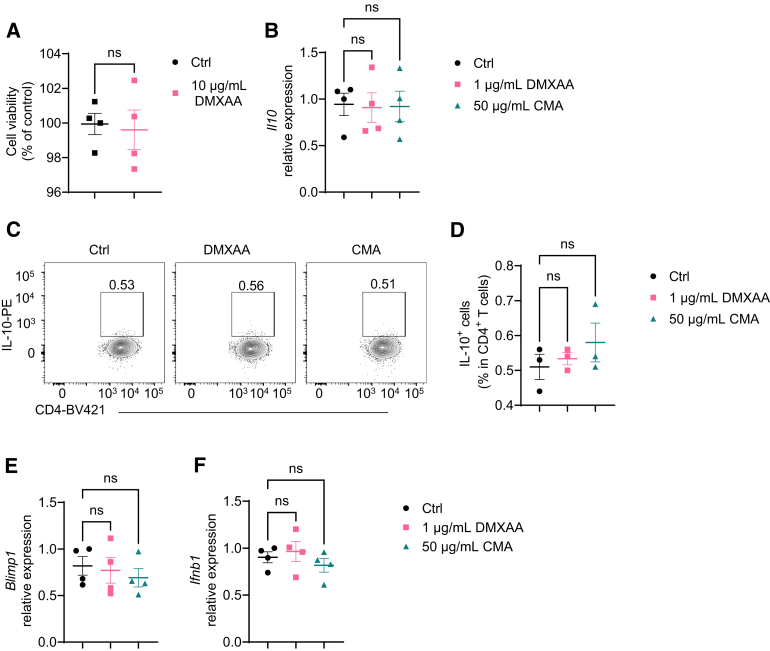
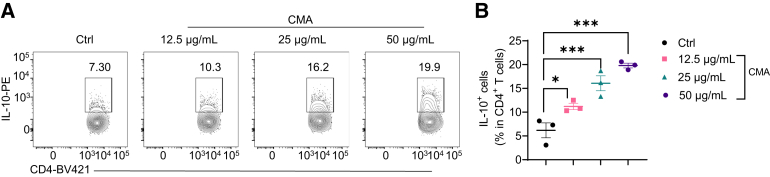
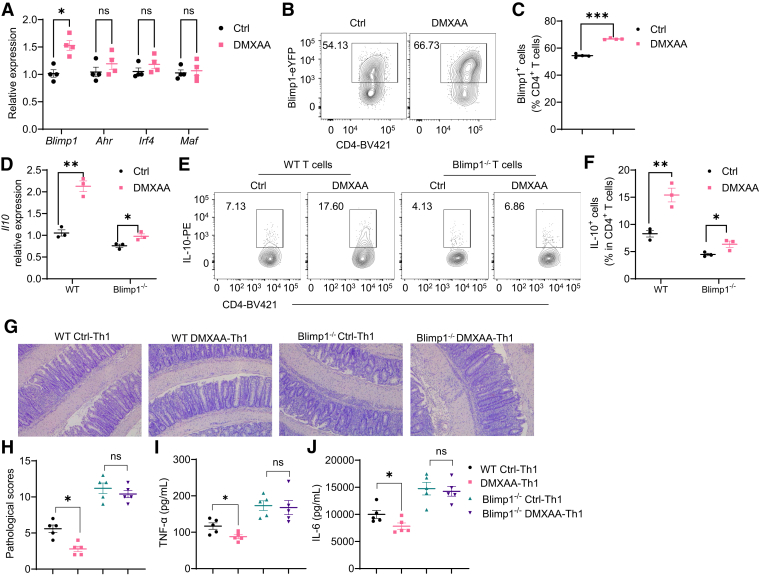
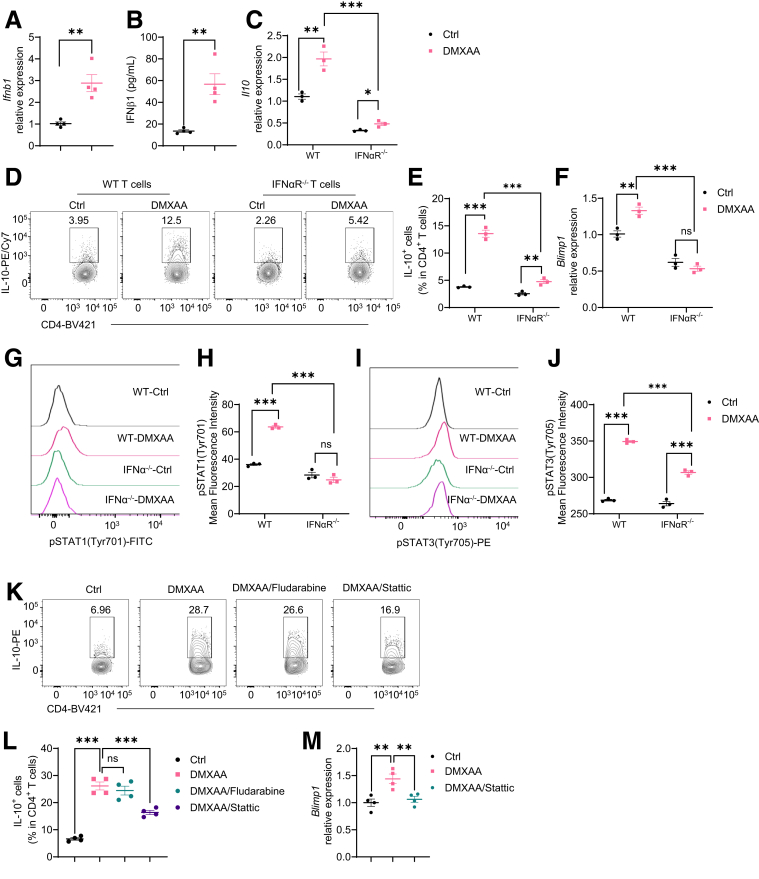
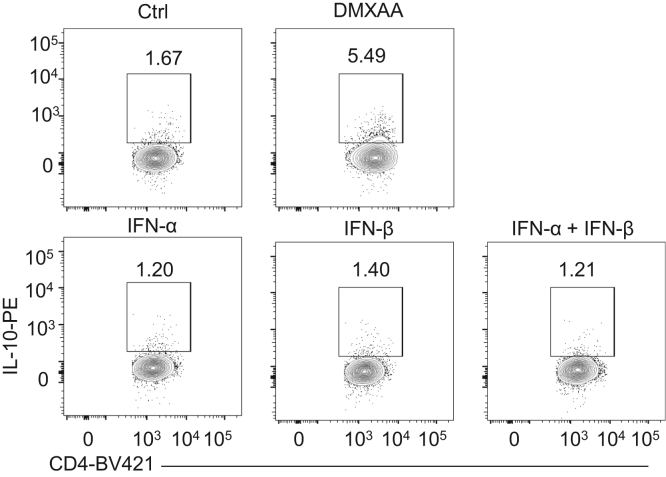
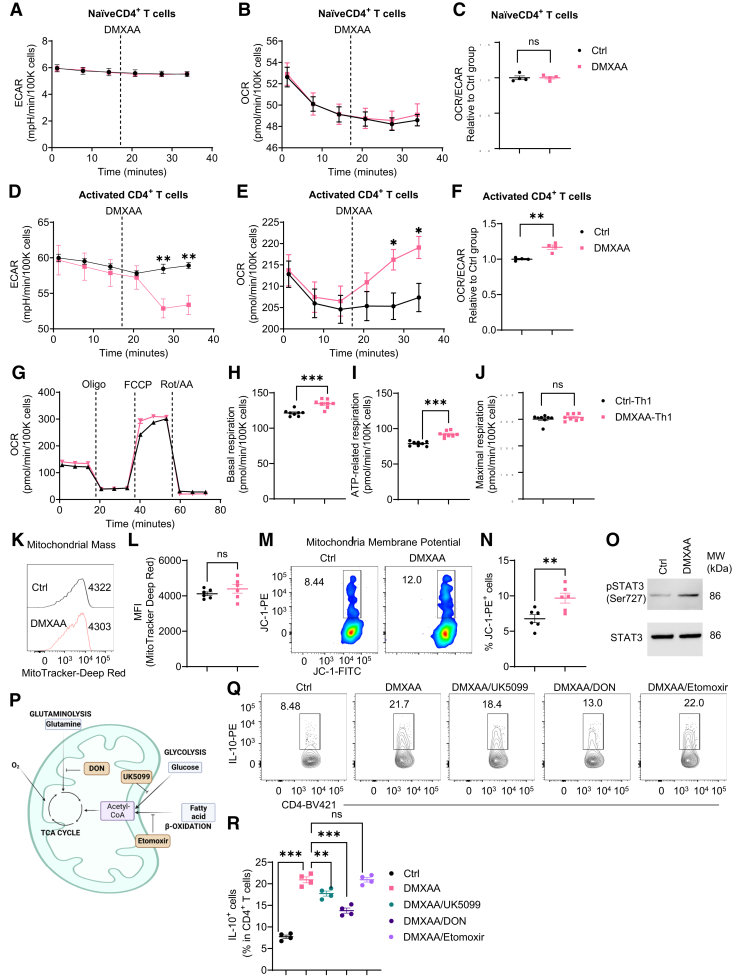
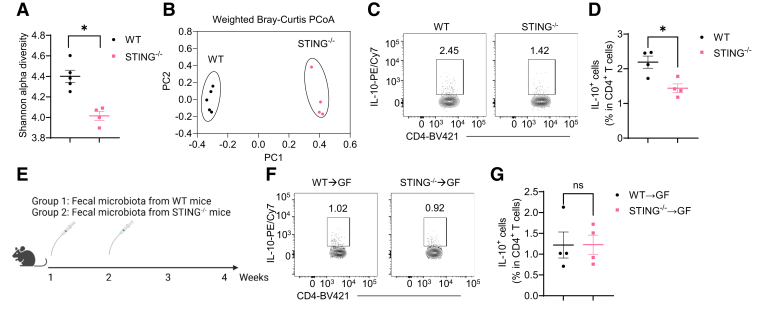
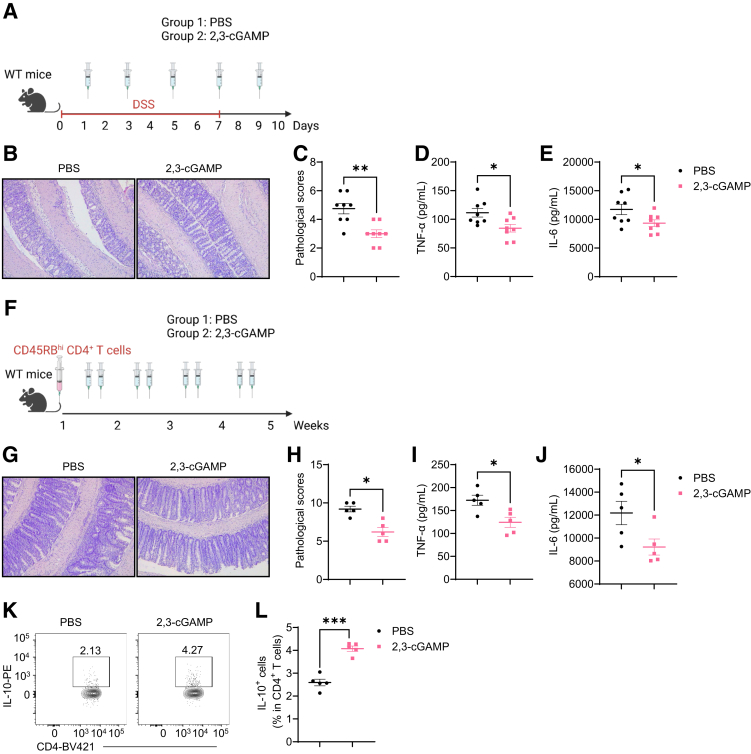
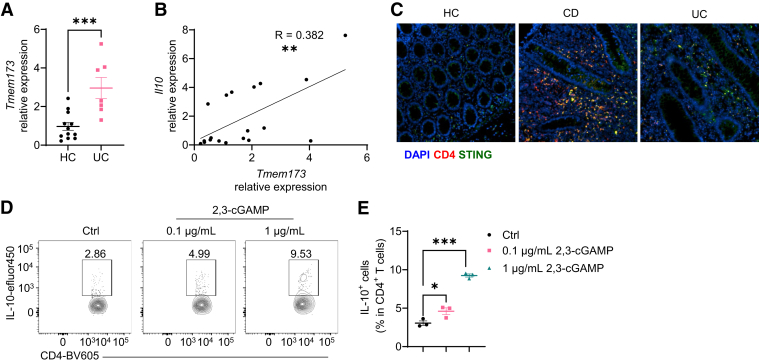
Results: Activation of STING transformed proinflammatory IFNγ+Th1 cells into IL-10+IFNγ+Th1 cells, which were dramatically less pathogenic in inducing colitis. STING promoted Th1 interleukin (IL)-10 production by inducing STAT3 translocation into nuclear and mitochondria, which promoted Blimp1 expression and mitochondrial oxidation, respectively. Blockade of glucose or glutamine-derived oxidation, but not lipid-derived oxidation, suppressed STING induction of IL-10. Gut microbiota were changed in STING-/- mice, but the altered microbiota did not mediate STING effects on intestinal CD4+T cell production of IL-10. Translationally, STING agonists suppressed both acute and chronic colitis. Intestinal STING+ CD4+T cells were increased in inflammatory bowel disease patients, and STING agonists upregulated IL-10 production in human CD4+T cells.
Conclusions: These findings establish a crucial role of T cell-intrinsic STING in switching off the pathogenic programs of Th1 cells in intestinal inflammation.
Keywords: Colitis; Mitochondrial Oxidation; STING; Th1 Cells.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
The STING That Tames Pro-inflammatory T-cells.Cell Mol Gastroenterol Hepatol. 2023;15(5):1251-1252. doi: 10.1016/j.jcmgh.2023.02.008. Epub 2023 Mar 6. Cell Mol Gastroenterol Hepatol. 2023. PMID: 36893791 Free PMC article. No abstract available.
References
-
- Lafaille J.J. The role of helper T cell subsets in autoimmune diseases. Cytokine Growth Factor Rev. 1998;9:139–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
