

A novel compound targets the feline infectious peritonitis virus nucleocapsid protein and inhibits viral replication in cell culture
- PMID: 36738790
- PMCID: PMC10011503
- DOI: 10.1016/j.jbc.2023.102976
A novel compound targets the feline infectious peritonitis virus nucleocapsid protein and inhibits viral replication in cell culture
Abstract
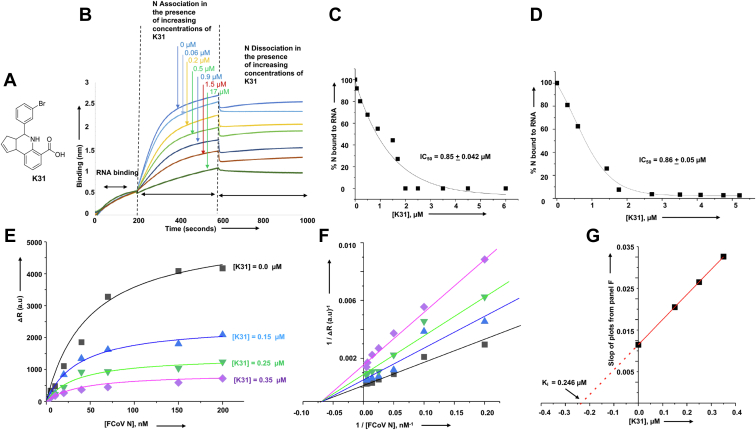
Feline infectious peritonitis (FIP) is a serious viral illness in cats, caused by feline coronavirus. Once a cat develops clinical FIP, the prognosis is poor. The effective treatment strategy for coronavirus infections with immunopathological complications such as SARS-CoV-2, MERS, and FIP is focused on antiviral and immunomodulatory agents to inhibit virus replication and enhance the protective immune response. In this article we report the binding and conformational alteration of feline alphacoronavirus (FCoV) nucleocapsid protein by a novel compound K31. K31 noncompetitively inhibited the interaction between the purified nucleocapsid protein and the synthetic 5' terminus of viral genomic RNA in vitro. K31 was well tolerated by cells and inhibited FCoV replication in cell culture with a selective index of 115. A single dose of K31inhibited FCoV replication to an undetectable level in 24 h post treatment. K31 did not affect the virus entry to the host cell but inhibited the postentry steps of virus replication. The nucleocapsid protein forms ribonucleocapsid in association with the viral genomic RNA that serves as a template for transcription and replication of the viral genome. Our results show that K31 treatment disrupted the structural integrity of ribonucleocapsid in virus-infected cells. After the COVID-19 pandemic, most of the antiviral drug development strategies have focused on RdRp and proteases encoded by the viral genome. Our results have shown that nucleocapsid protein is a druggable target for anticoronavirus drug discovery.
Keywords: antivirals; coronavirus; nucleocapsid protein; virus; virus replication.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures








References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
