

Pivotal role for S-nitrosylation of DNA methyltransferase 3B in epigenetic regulation of tumorigenesis
- PMID: 36739439
- PMCID: PMC9899281
- DOI: 10.1038/s41467-023-36232-6
Pivotal role for S-nitrosylation of DNA methyltransferase 3B in epigenetic regulation of tumorigenesis
Abstract
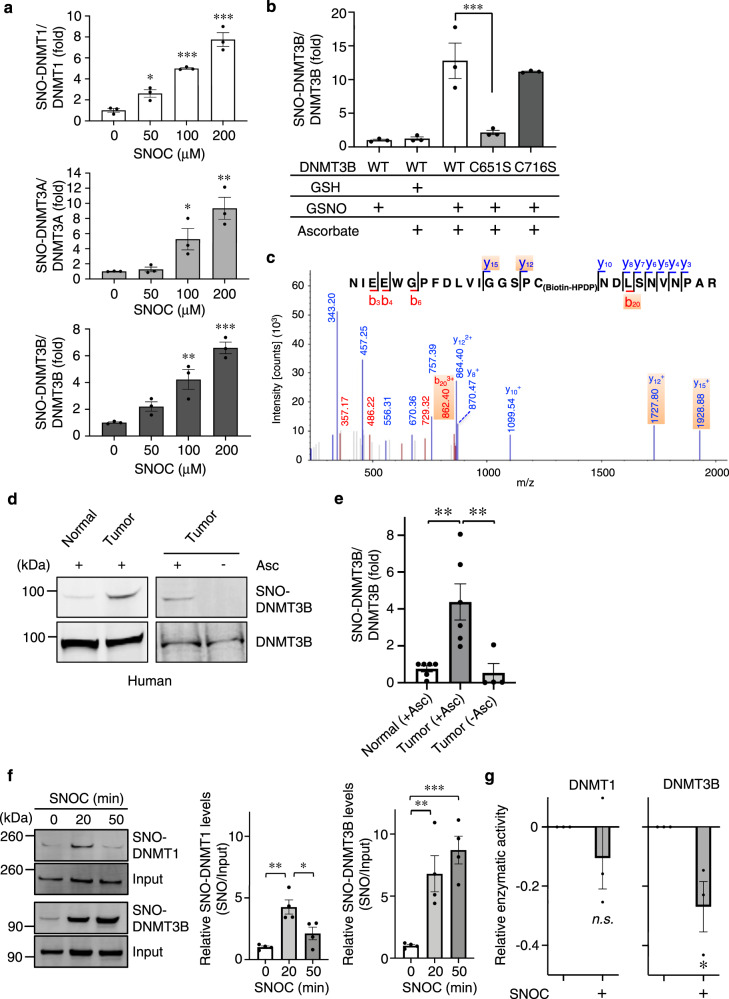
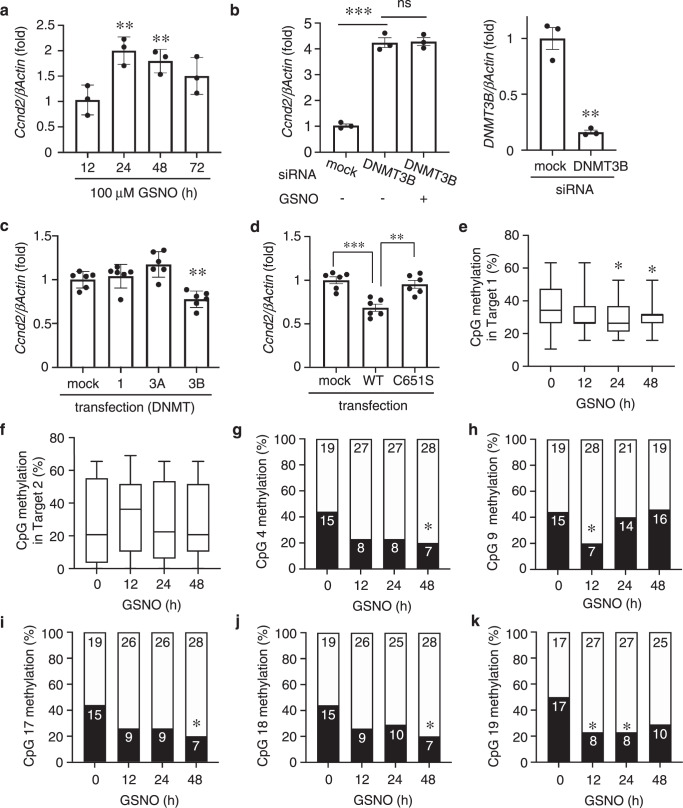
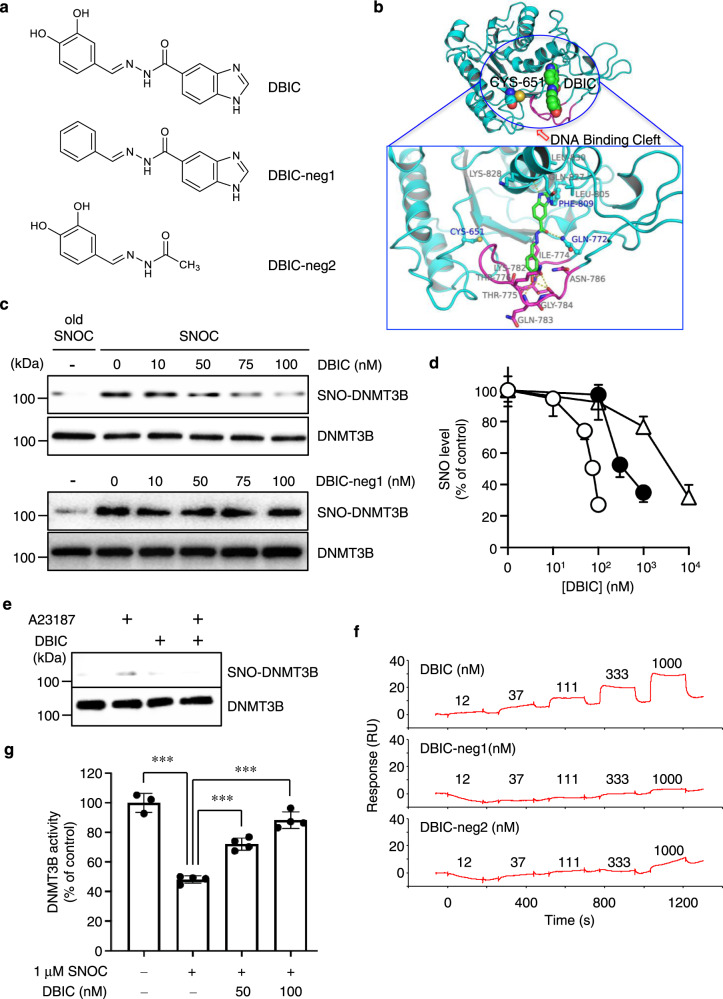
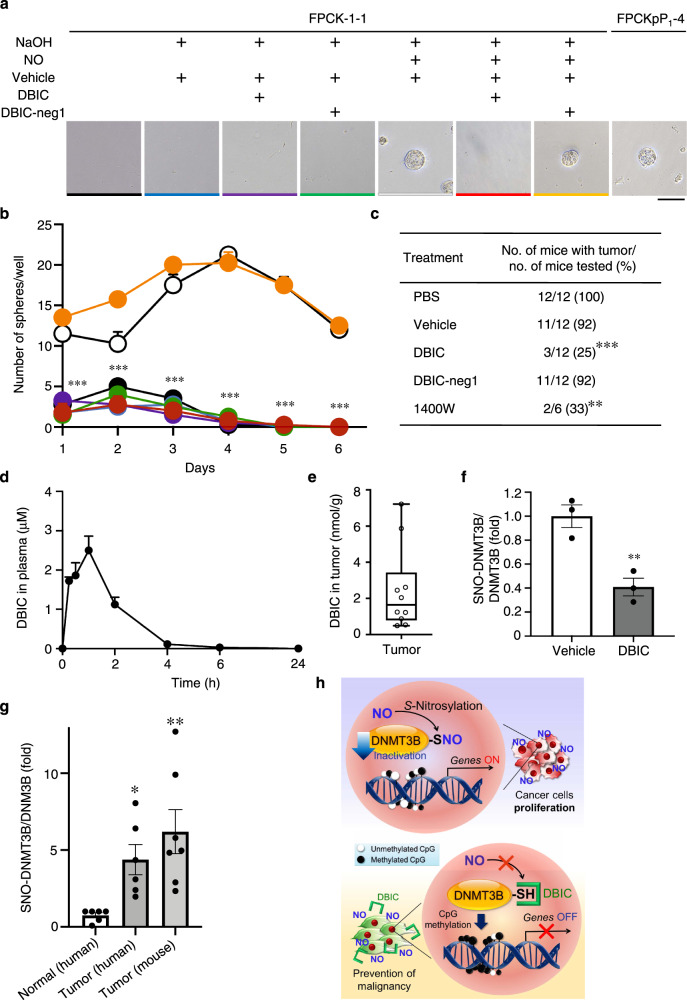
DNA methyltransferases (DNMTs) catalyze methylation at the C5 position of cytosine with S-adenosyl-L-methionine. Methylation regulates gene expression, serving a variety of physiological and pathophysiological roles. The chemical mechanisms regulating DNMT enzymatic activity, however, are not fully elucidated. Here, we show that protein S-nitrosylation of a cysteine residue in DNMT3B attenuates DNMT3B enzymatic activity and consequent aberrant upregulation of gene expression. These genes include Cyclin D2 (Ccnd2), which is required for neoplastic cell proliferation in some tumor types. In cell-based and in vivo cancer models, only DNMT3B enzymatic activity, and not DNMT1 or DNMT3A, affects Ccnd2 expression. Using structure-based virtual screening, we discovered chemical compounds that specifically inhibit S-nitrosylation without directly affecting DNMT3B enzymatic activity. The lead compound, designated DBIC, inhibits S-nitrosylation of DNMT3B at low concentrations (IC50 ≤ 100 nM). Treatment with DBIC prevents nitric oxide (NO)-induced conversion of human colonic adenoma to adenocarcinoma in vitro. Additionally, in vivo treatment with DBIC strongly attenuates tumor development in a mouse model of carcinogenesis triggered by inflammation-induced generation of NO. Our results demonstrate that de novo DNA methylation mediated by DNMT3B is regulated by NO, and DBIC protects against tumor formation by preventing aberrant S-nitrosylation of DNMT3B.
© 2023. The Author(s).
Conflict of interest statement
T. Uehara, A.I., A.K., K.Y.J.Z., and F.O. are inventors on a patent (JP6887640) related to the effects of specific inhibitors of
Figures
References
-
- Lipton SA, et al. A redox-based mechanism for the neuroprotective and neurodestructive effects of nitric oxide and related nitroso-compounds. Nature. 1993;364:626–632. - PubMed
-
- Stamler JS. Redox signaling: nitrosylation and related target interactions of nitric oxide. Cell. 1994;78:931–936. - PubMed
-
- Wolhuter K, Eaton P. How widespread is stable protein S-nitrosylation as an end-effector of protein regulation? Free Radic. Biol. Med. 2017;109:156–166. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
