

SOX9 Modulates the Transformation of Gastric Stem Cells Through Biased Symmetric Cell Division
- PMID: 36740200
- PMCID: PMC10200757
- DOI: 10.1053/j.gastro.2023.01.037
SOX9 Modulates the Transformation of Gastric Stem Cells Through Biased Symmetric Cell Division
Abstract
Background & aims: Transformation of stem/progenitor cells has been associated with tumorigenesis in multiple tissues, but stem cells in the stomach have been hard to localize. We therefore aimed to use a combination of several markers to better target oncogenes to gastric stem cells and understand their behavior in the initial stages of gastric tumorigenesis.
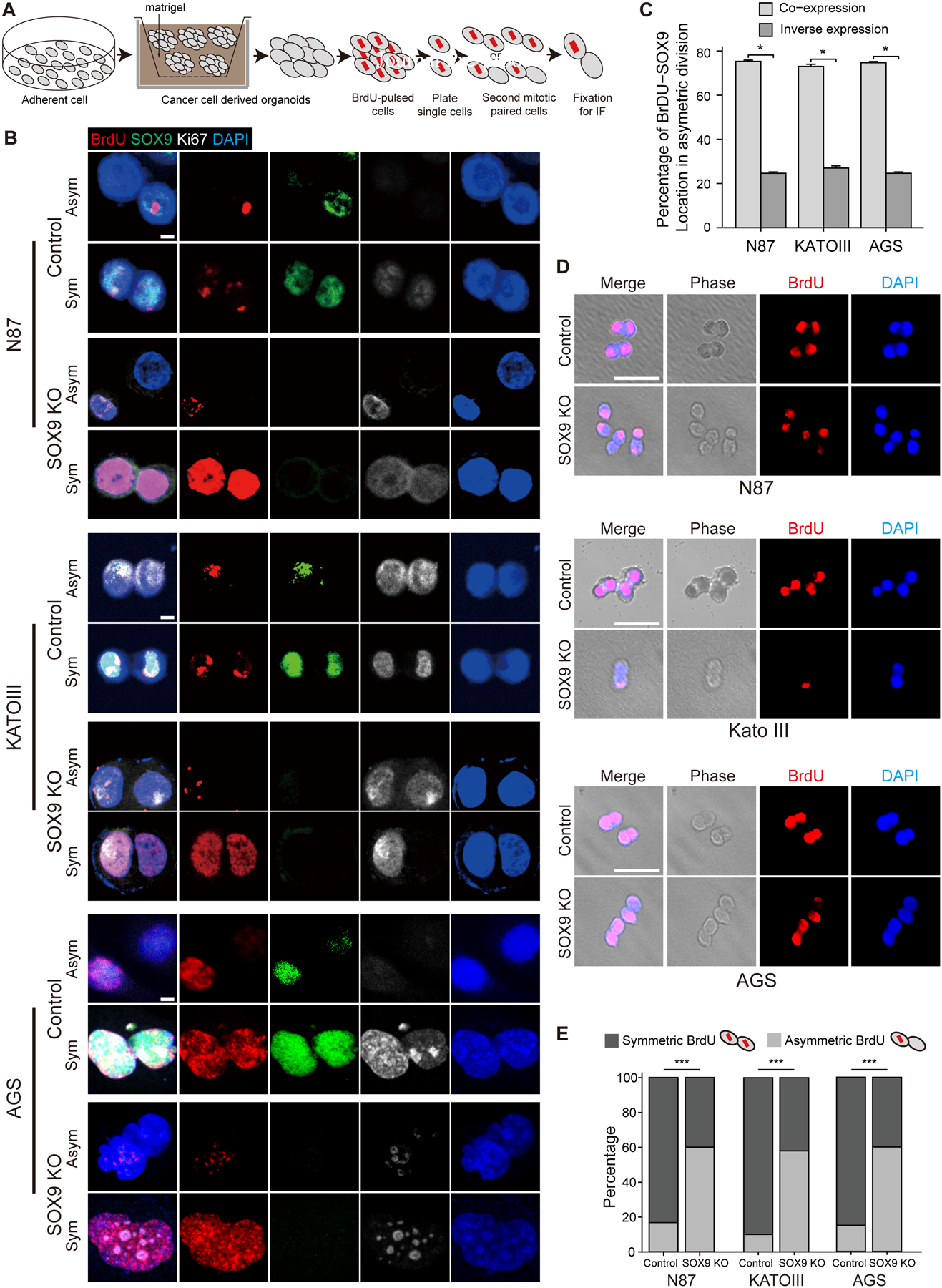
Methods: Mouse models of gastric metaplasia and cancer by targeting stem/progenitor cells were generated and analyzed with techniques including reanalysis of single-cell RNA sequencing and immunostaining. Gastric cancer cell organoids were genetically manipulated with clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) for functional studies. Cell division was determined by bromodeoxyuridine-chasing assay and the assessment of the orientation of the mitotic spindles. Gastric tissues from patients were examined by histopathology and immunostaining.
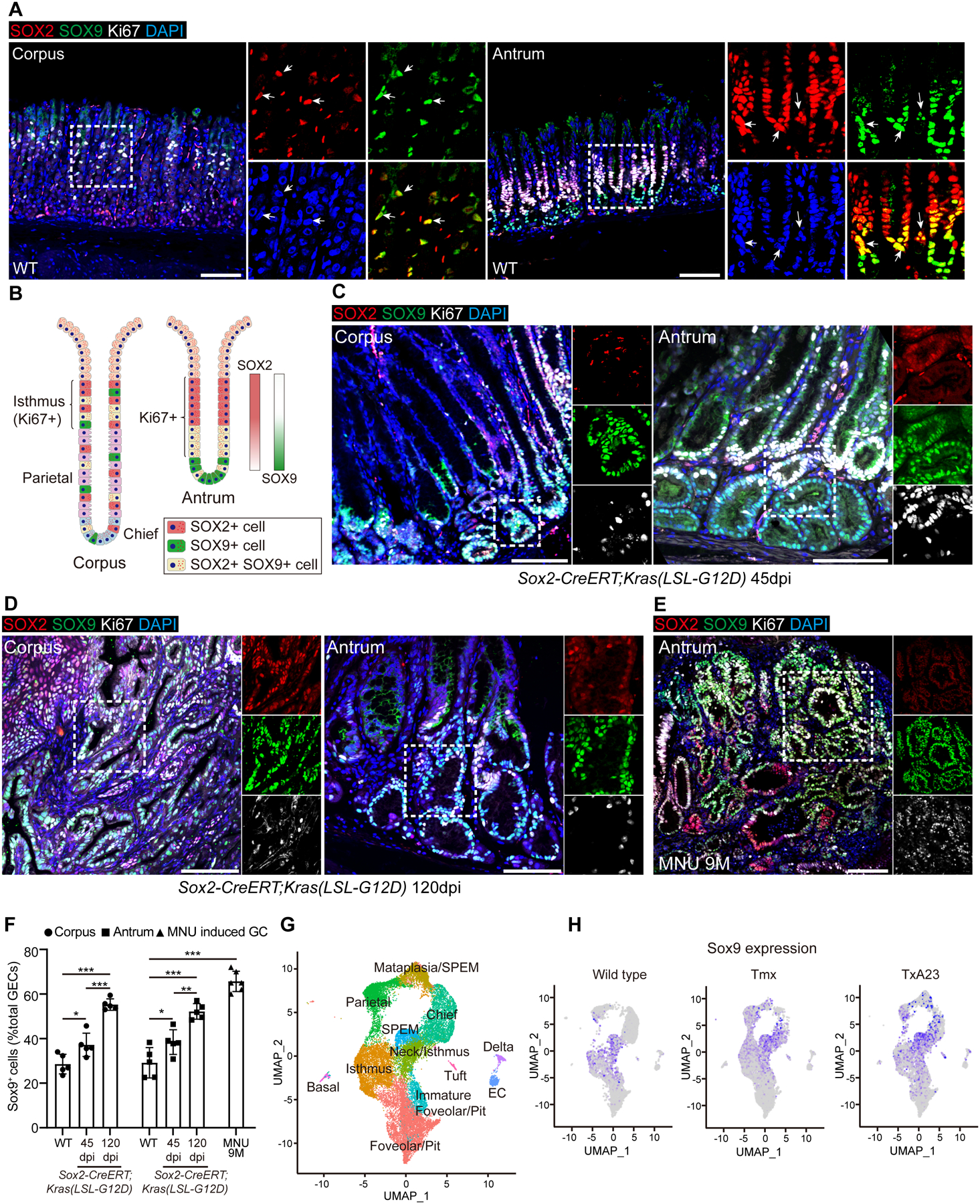
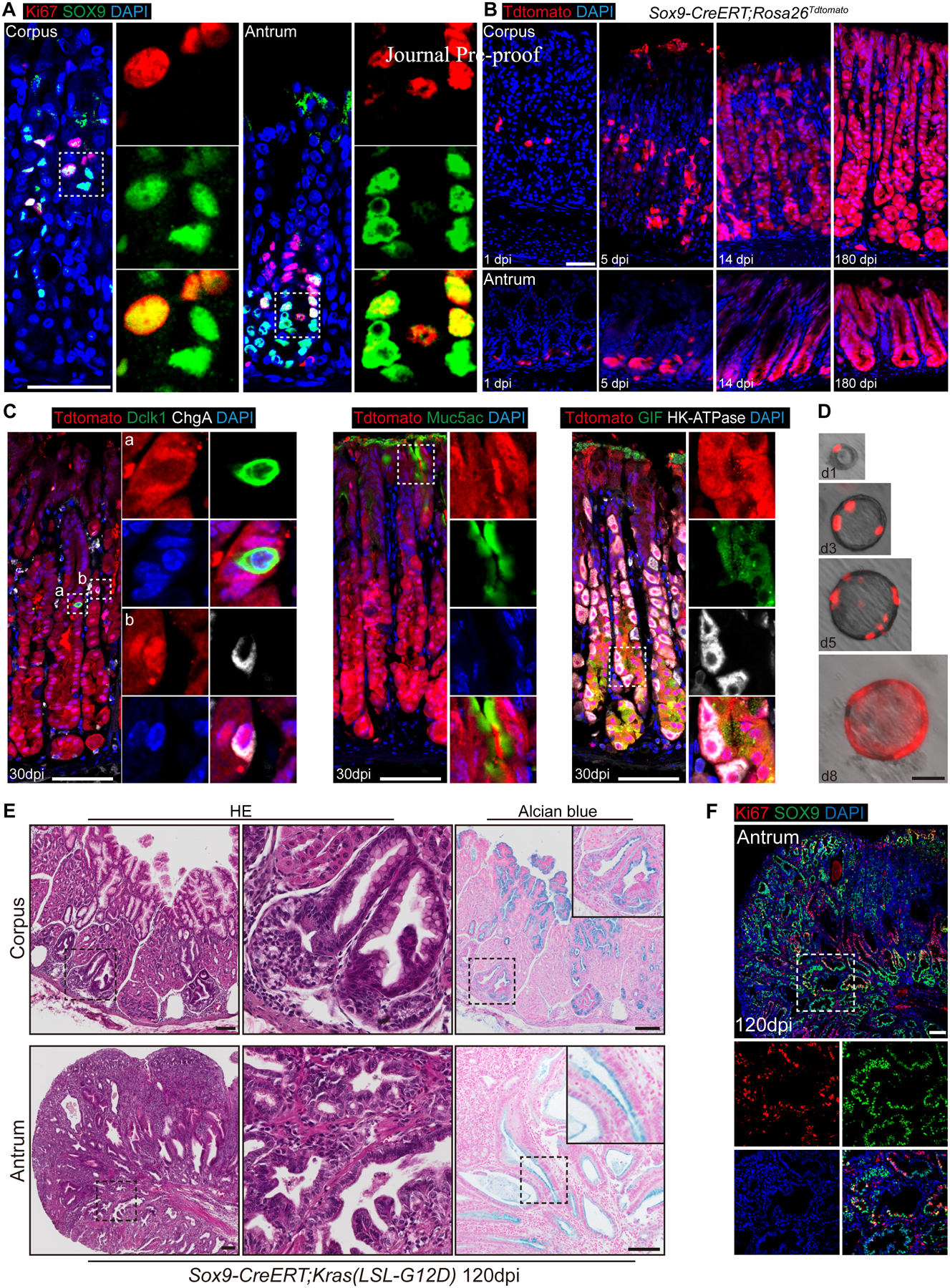
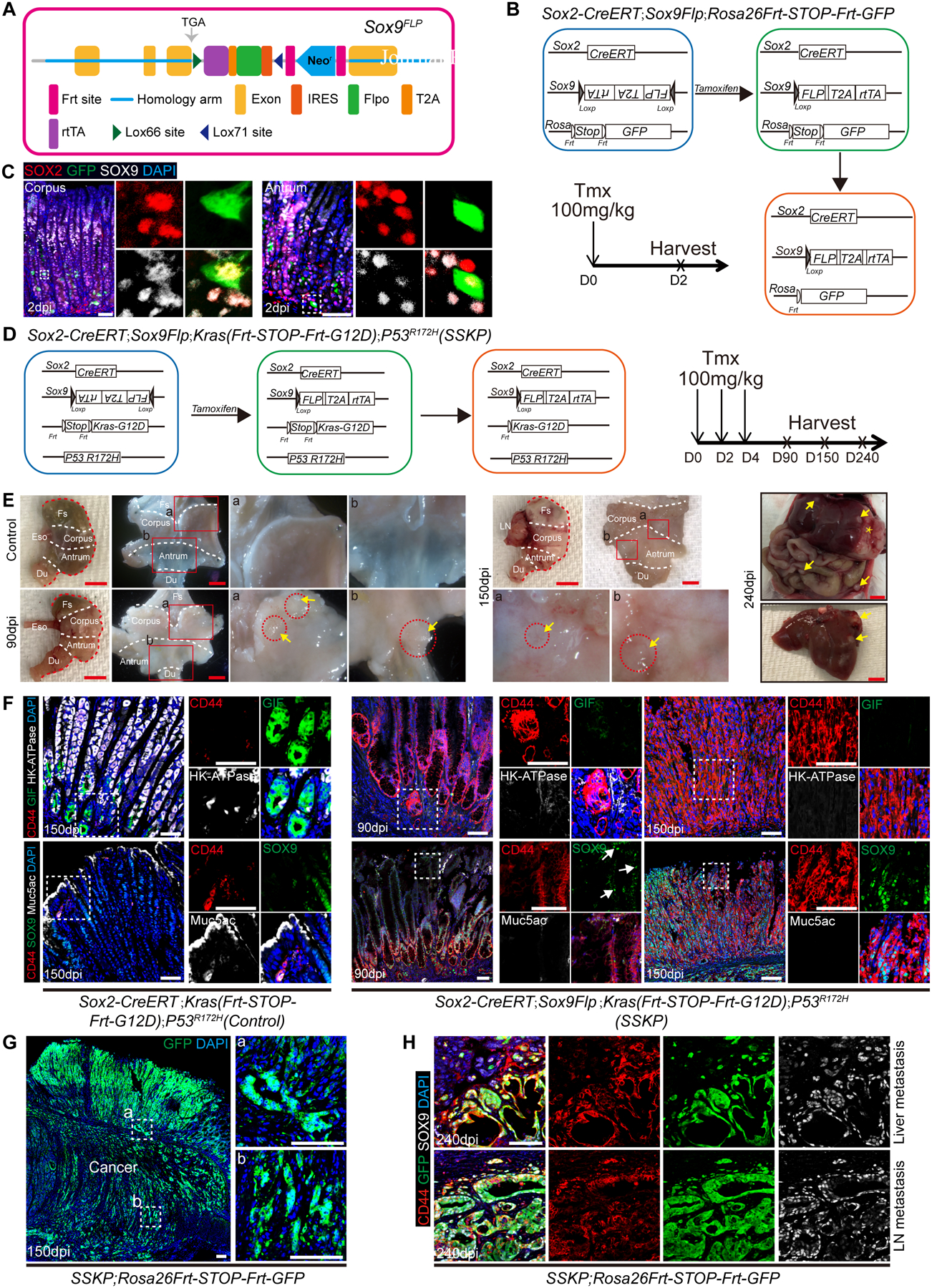
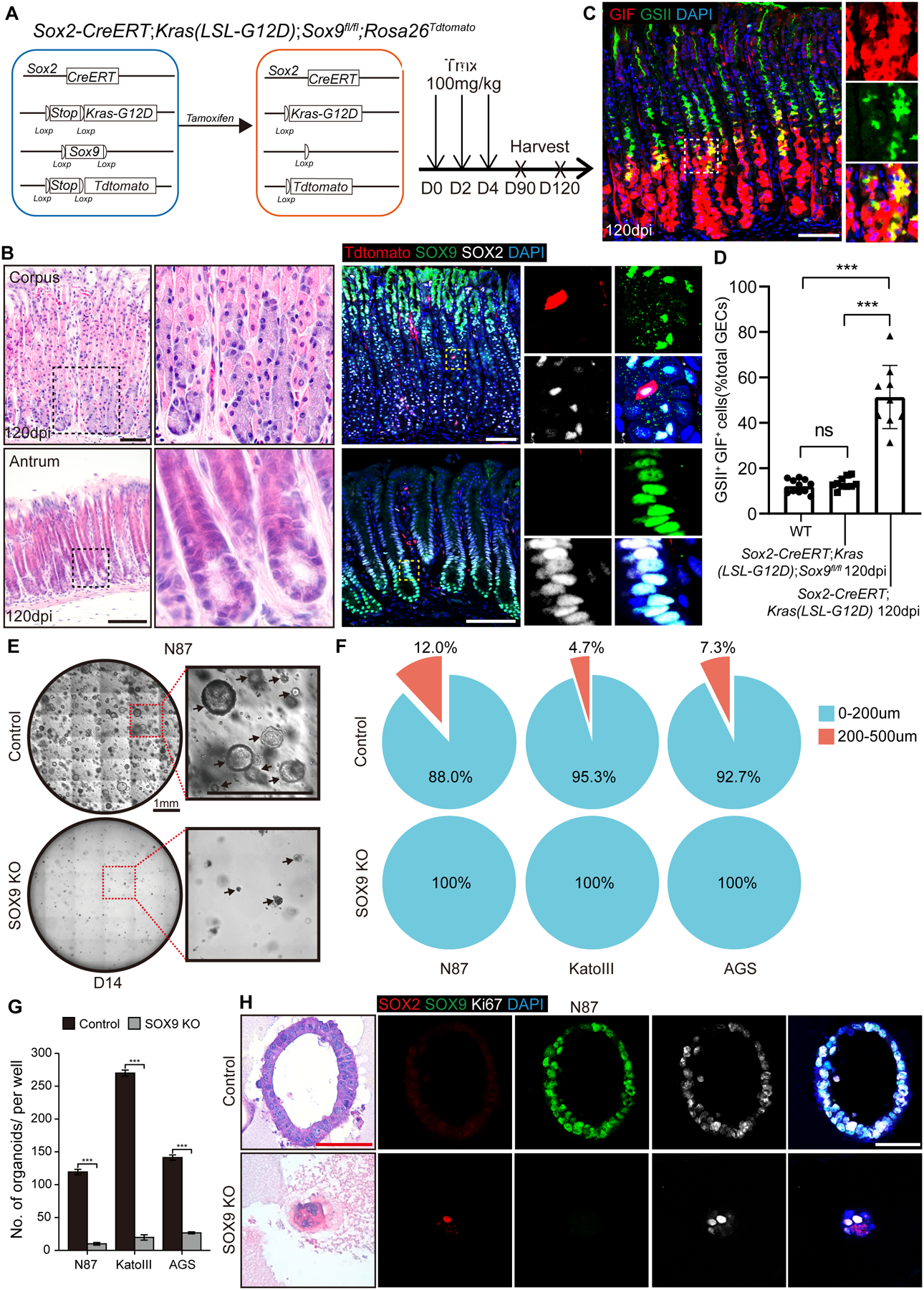
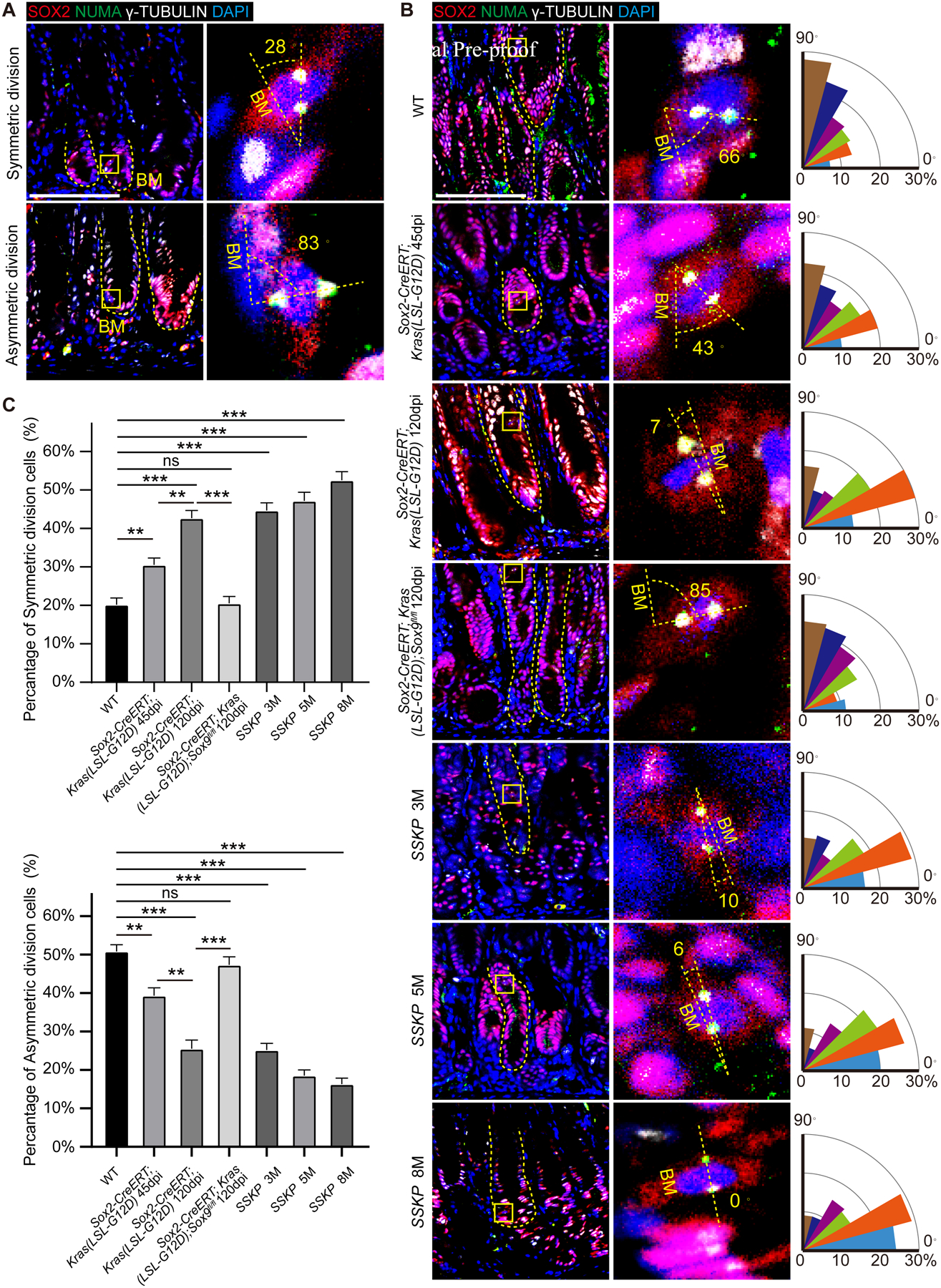
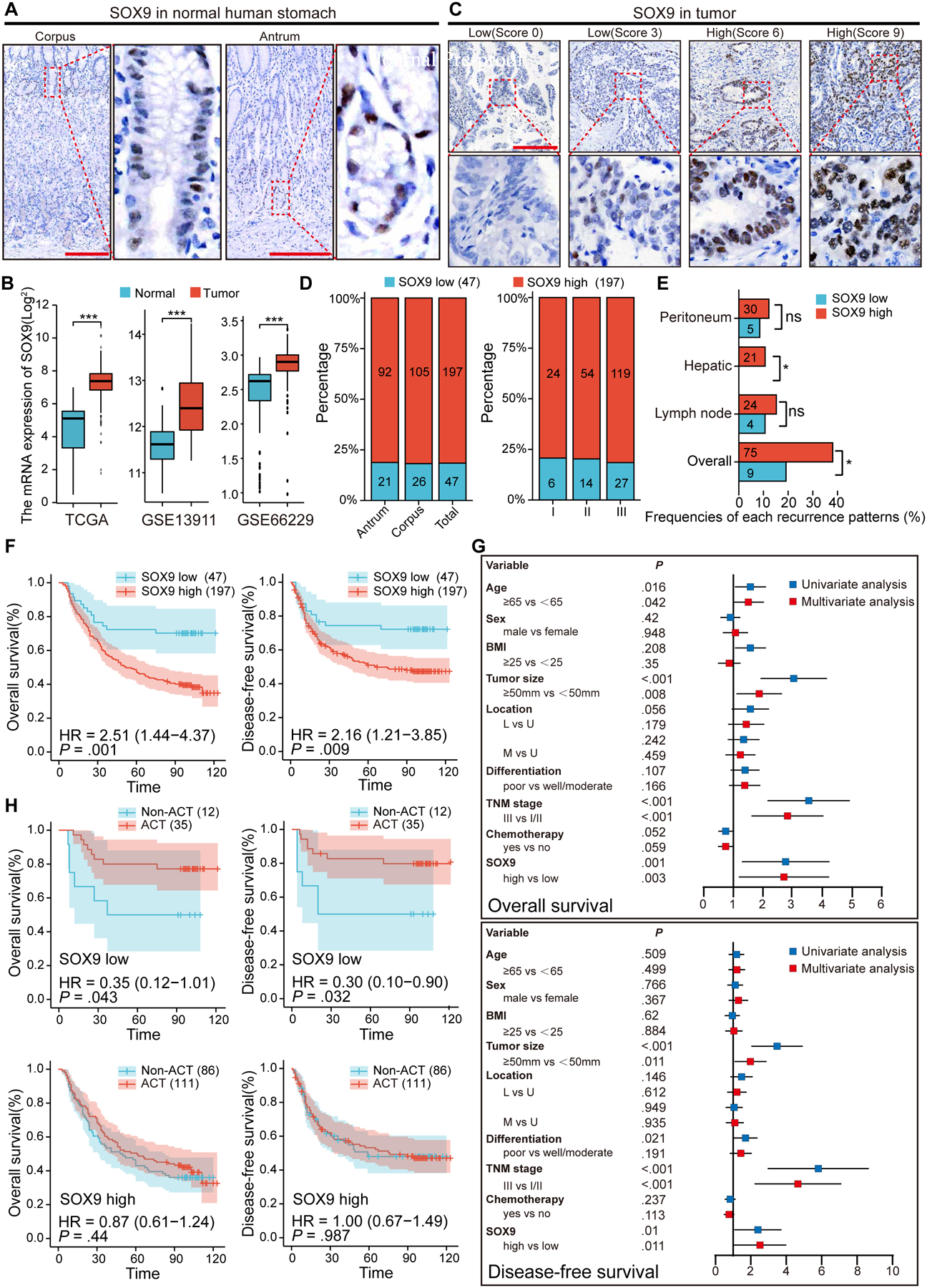
Results: Oncogenic insults lead to expansion of SOX9+ progenitor cells in the mouse stomach. Genetic lineage tracing and organoid culture studies show that SOX9+ gastric epithelial cells overlap with SOX2+ progenitors and include stem cells that can self-renew and differentiate to generate all gastric epithelial cells. Moreover, oncogenic targeting of SOX9+SOX2+ cells leads to invasive gastric cancer in our novel mouse model (Sox2-CreERT;Sox9-loxp(66)-rtTA-T2A-Flpo-IRES-loxp(71);Kras(Frt-STOP-Frt-G12D);P53R172H), which combines Cre-loxp and Flippase-Frt genetic recombination systems. Sox9 deletion impedes the expansion of gastric progenitor cells and blocks neoplasia after Kras activation. Although Sox9 is not required for maintaining tissue homeostasis where asymmetric division predominates, loss of Sox9 in the setting of Kras activation leads to reduced symmetric cell division and effectively attenuates the Kras-dependent expansion of stem/progenitor cells. Similarly, Sox9 deletion in gastric cancer organoids reduces symmetric cell division, organoid number, and organoid size. In patients with gastric cancer, high levels of SOX9 are associated with recurrence and poor prognosis.
Conclusion: SOX9 marks gastric stem cells and modulates biased symmetric cell division, which appears to be required for the malignant transformation of gastric stem cells.
Keywords: Gastric Cancer; Metaplasia; Mouse Model; Neoplasia; SOX2.
Copyright © 2023 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
Comment in
-
Sox9, A Key Regulator of Gastric Stem Cell Behavior Driving Cancer Initiation.Gastroenterology. 2023 Jun;164(7):1052-1053. doi: 10.1053/j.gastro.2023.03.215. Epub 2023 Mar 24. Gastroenterology. 2023. PMID: 36965742 No abstract available.
-
SOX9 Expression Pattern and Its Prognosis in Gastric Cancer Should be Fully Evaluated.Gastroenterology. 2023 Oct;165(4):1094. doi: 10.1053/j.gastro.2023.06.024. Epub 2023 Jul 4. Gastroenterology. 2023. PMID: 37414194 No abstract available.
References
-
- Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a cancer journal for clinicians 2021;71:209–249. - PubMed
-
- Barker N, Ridgway R, van Es J, et al. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 2009;457:608–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10 RR027050/RR/NCRR NIH HHS/United States
- R01 CA266528/CA/NCI NIH HHS/United States
- R01 DK100342/DK/NIDDK NIH HHS/United States
- R01 CA272901/CA/NCI NIH HHS/United States
- P30 DK132710/DK/NIDDK NIH HHS/United States
- S10 OD020056/OD/NIH HHS/United States
- P30 CA013696/CA/NCI NIH HHS/United States
- R01 DK120650/DK/NIDDK NIH HHS/United States
- IK6 BX006300/BX/BLRD VA/United States
- I01 BX002115/BX/BLRD VA/United States
- P01 CA268991/CA/NCI NIH HHS/United States
- R01 DK113144/DK/NIDDK NIH HHS/United States
- R01 CA138833/CA/NCI NIH HHS/United States
- R01 DK132251/DK/NIDDK NIH HHS/United States
- R01 DK114436/DK/NIDDK NIH HHS/United States
- R01 CA249949/CA/NCI NIH HHS/United States
- P01 CA098101/CA/NCI NIH HHS/United States
- S10 OD032447/OD/NIH HHS/United States
- IK6 BX003787/BX/BLRD VA/United States
- I01 BX001179/BX/BLRD VA/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
