

Limited Proteolysis-Mass Spectrometry Reveals Aging-Associated Changes in Cerebrospinal Fluid Protein Abundances and Structures
- PMID: 36741774
- PMCID: PMC9893943
- DOI: 10.1038/s43587-022-00196-x
Limited Proteolysis-Mass Spectrometry Reveals Aging-Associated Changes in Cerebrospinal Fluid Protein Abundances and Structures
Erratum in
-
Publisher Correction: Limited proteolysis-mass spectrometry reveals aging-associated changes in cerebrospinal fluid protein abundances and structures.Nat Aging. 2022 May;2(5):455. doi: 10.1038/s43587-022-00225-9. Nat Aging. 2022. PMID: 37118077 No abstract available.
Abstract
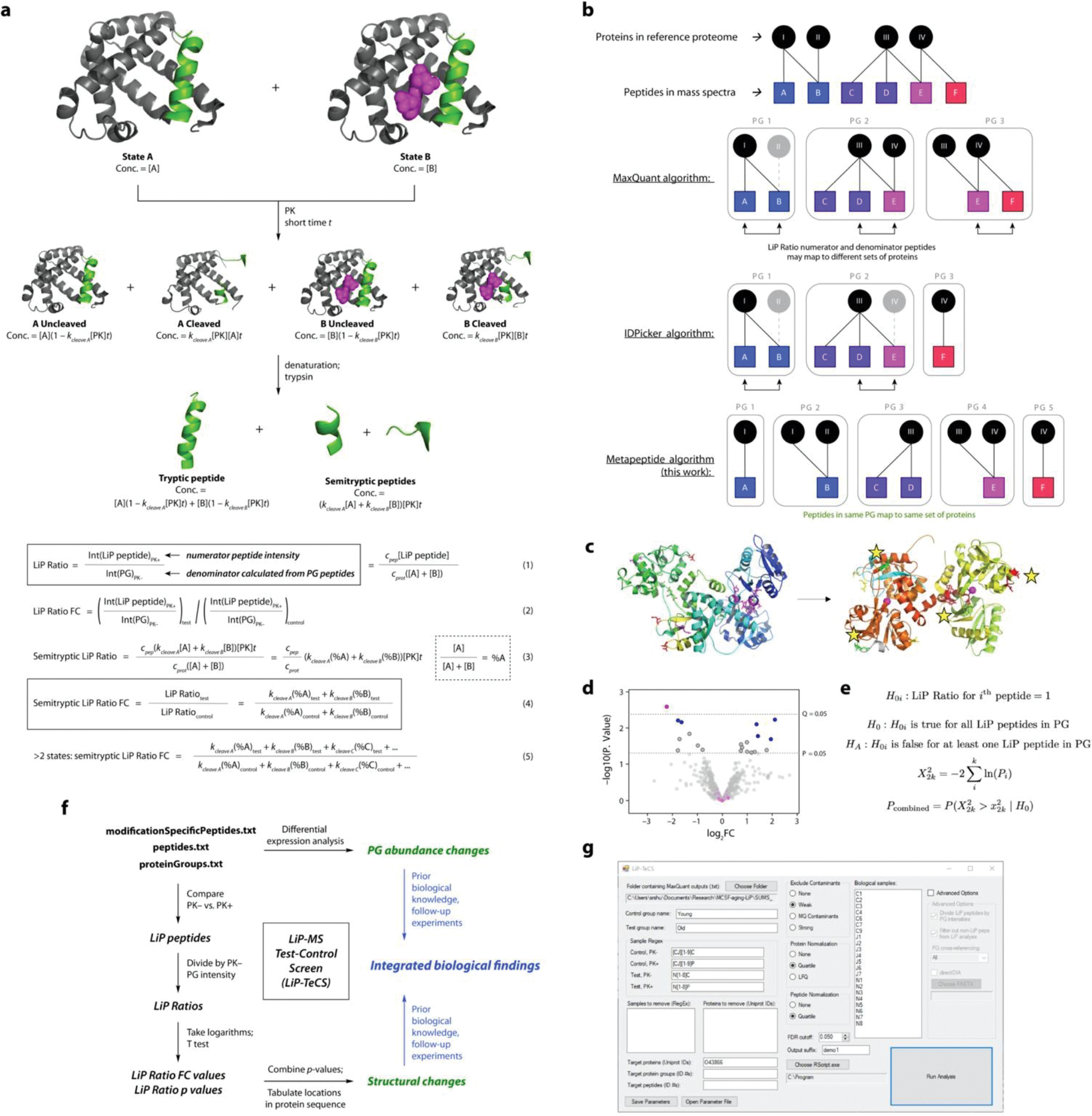
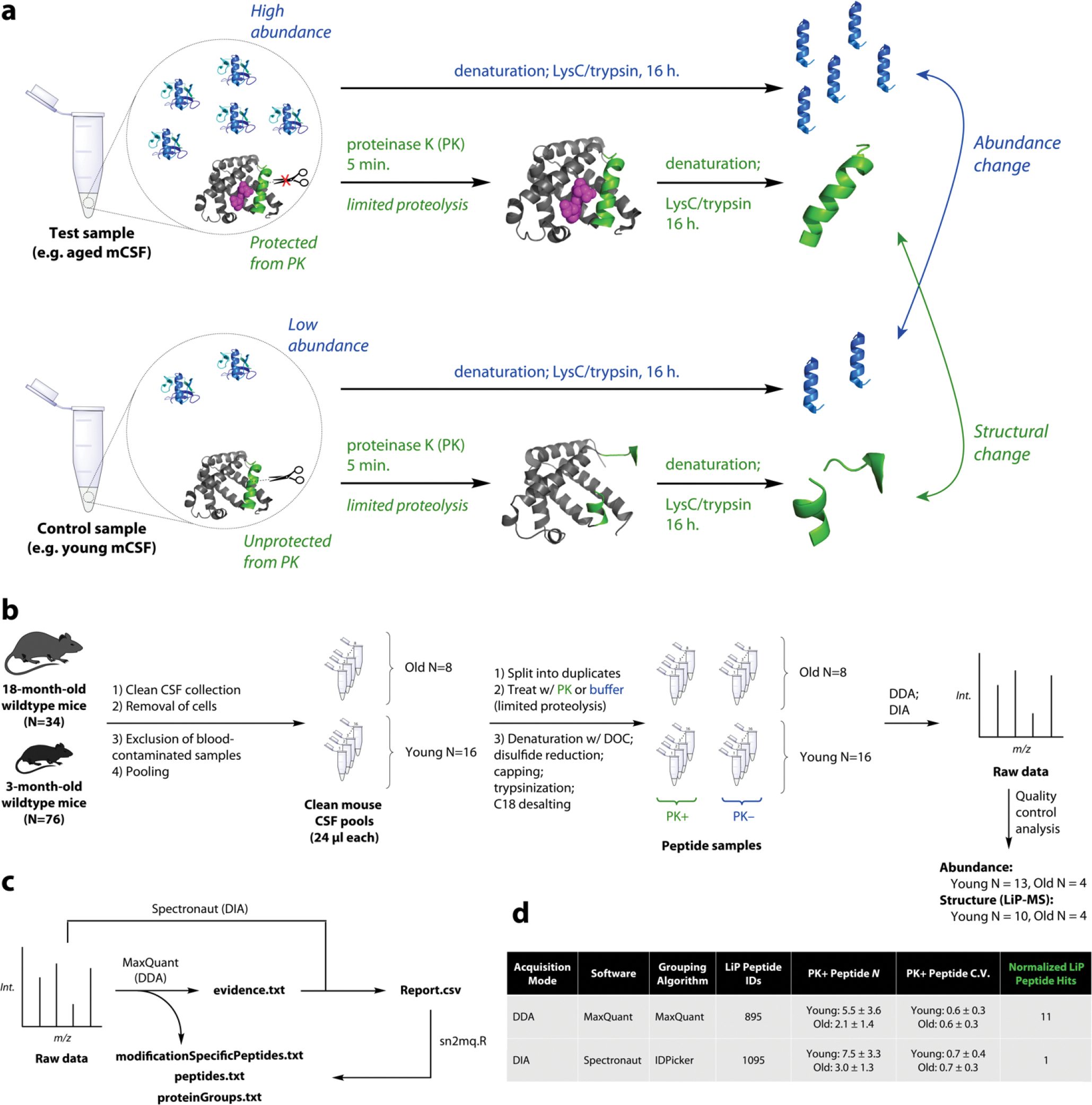
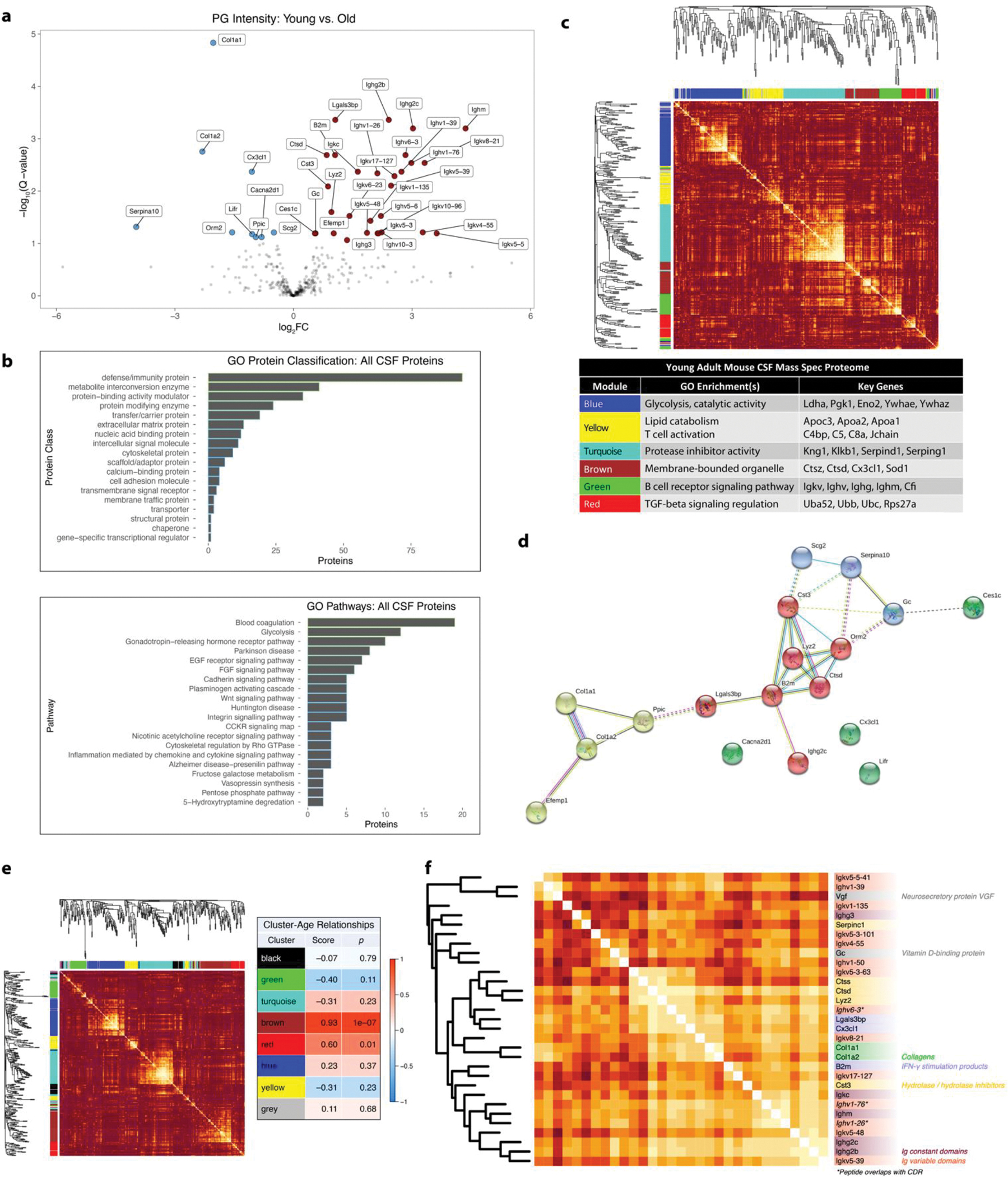
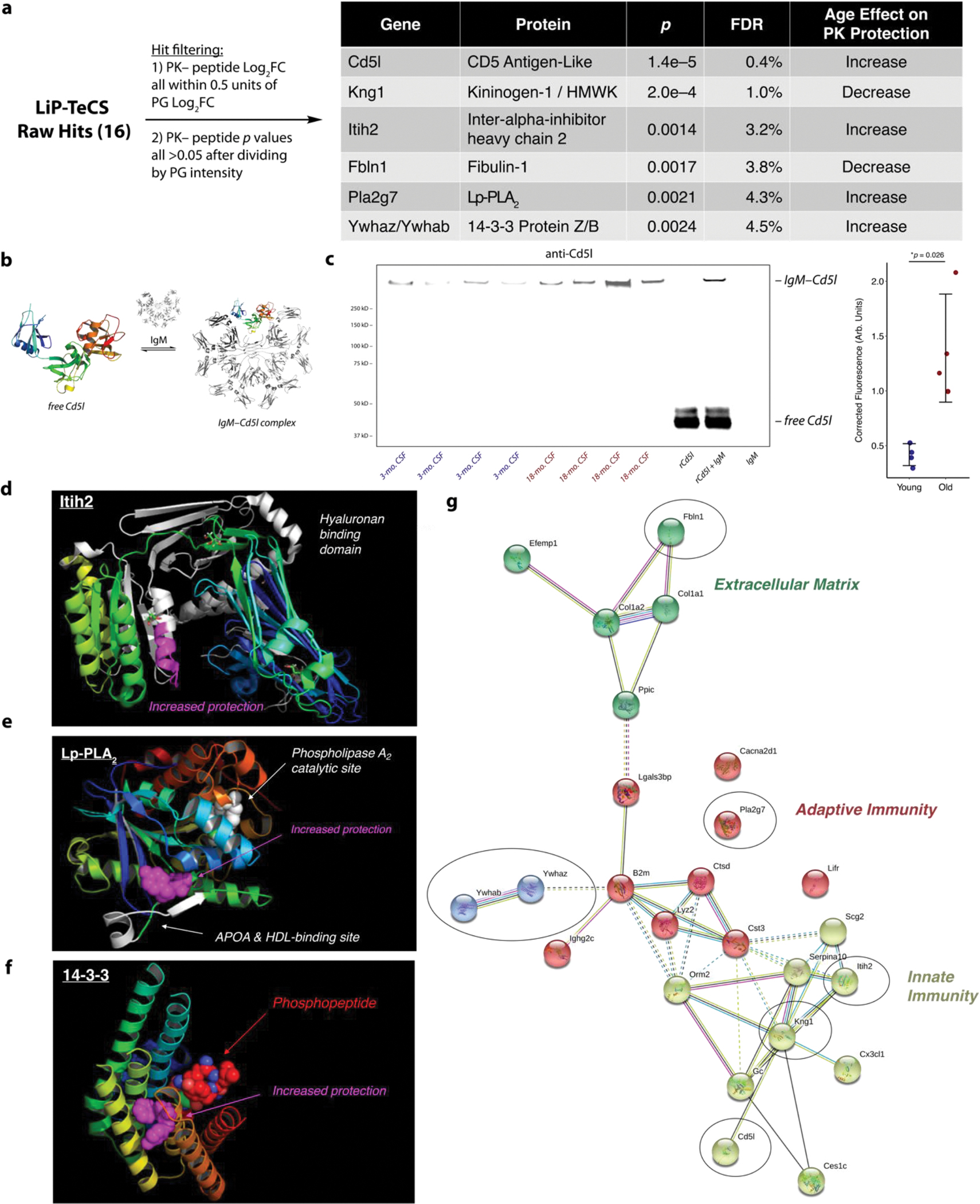
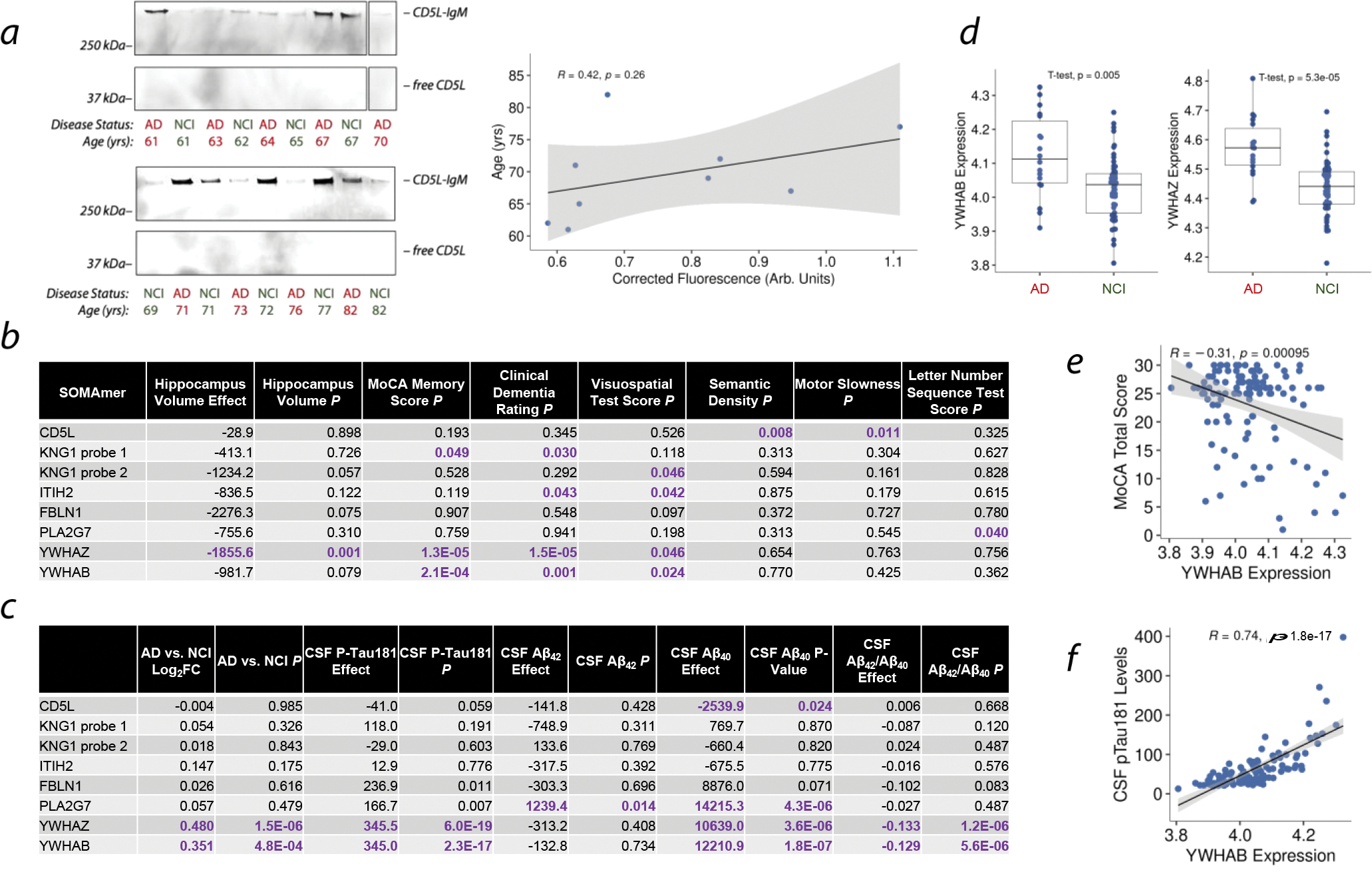
Cerebrospinal fluid (CSF) proteins and their structures have been implicated repeatedly in aging and neurodegenerative diseases. Limited proteolysis-mass spectrometry (LiP-MS) is a method that enables proteome-wide screening for changes in both protein abundance and structure. To screen for novel aging-associated changes in the CSF proteome, we performed LiP-MS on CSF from young and old mice with a modified analysis pipeline. We found 38 protein groups change in abundance with aging, most dominantly immunoglobulins of the IgM subclass. We discovered six high-confidence candidates that appeared to change in structure with aging, of which Kng1, Itih2, Lp-PLA2, and 14-3-3 proteins have binding partners or proteoforms known previously to change in the brain with Alzheimer's disease. Intriguingly, using orthogonal validation by Western blot we found the LiP-MS hit Cd5l forms a covalent complex with IgM in mouse and human CSF whose abundance increases with aging. SOMAmer probe signals for all six LiP-MS hits in human CSF, especially 14-3-3 proteins, significantly associate with several clinical features relevant to cognitive function and neurodegeneration. Together, our findings show that LiP-MS can uncover age-related structural changes in CSF with relevance to neurodegeneration.
Conflict of interest statement
Competing Interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
