

microRNA production in Arabidopsis
- PMID: 36743500
- PMCID: PMC9893293
- DOI: 10.3389/fpls.2023.1096772
microRNA production in Arabidopsis
Abstract
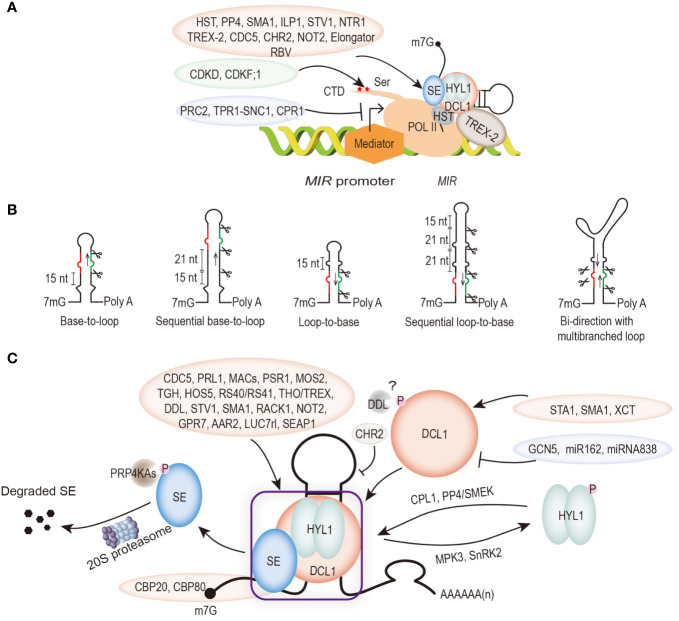
In plants, microRNAs (miRNAs) associate with ARGONAUTE (AGO) proteins and act as sequence-specific repressors of target gene expression, at the post-transcriptional level through target transcript cleavage and/or translational inhibition. MiRNAs are mainly transcribed by DNA-dependent RNA polymerase II (POL II) and processed by DICER LIKE1 (DCL1) complex into 21∼22 nucleotide (nt) long. Although the main molecular framework of miRNA biogenesis and modes of action have been established, there are still new requirements continually emerging in the recent years. The studies on the involvement factors in miRNA biogenesis indicate that miRNA biogenesis is not accomplished separately step by step, but is closely linked and dynamically regulated with each other. In this article, we will summarize the current knowledge on miRNA biogenesis, including MIR gene transcription, primary miRNA (pri-miRNA) processing, miRNA AGO1 loading and nuclear export; and miRNA metabolism including methylation, uridylation and turnover. We will describe how miRNAs are produced and how the different steps are regulated. We hope to raise awareness that the linkage between different steps and the subcellular regulation are becoming important for the understanding of plant miRNA biogenesis and modes of action.
Keywords: AGO; Arabidopsis; DCL; miRNA biogenesis; microprocessors.
Copyright © 2023 Ding and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Baranauskė S., Mickutė M., Plotnikova A., Finke A., Venclovas Č., Klimašauskas S., et al. (2015). Functional mapping of the plant small RNA methyltransferase: HEN1 physically interacts with HYL1 and DICER-LIKE 1 proteins. Nucleic Acids Res. 43, 2802–2812. doi: 10.1093/nar/gkv102 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials