

The binding pocket properties were fundamental to functional diversification of the GDSL-type esterases/lipases gene family in cotton
- PMID: 36743561
- PMCID: PMC9889996
- DOI: 10.3389/fpls.2022.1099673
The binding pocket properties were fundamental to functional diversification of the GDSL-type esterases/lipases gene family in cotton
Abstract
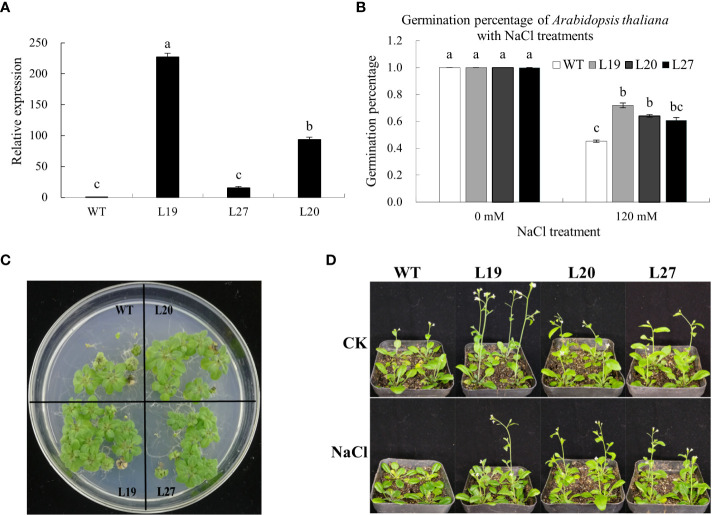
Cotton is one of the most important crops in the world. GDSL-type esterases/lipases (GELPs) are widely present in all kingdoms and play an essential role in regulating plant growth, development, and responses to abiotic and biotic stresses. However, the molecular mechanisms underlying this functional diversity remain unclear. Here, based on the identification of the GELP gene family, we applied genetic evolution and molecular simulation techniques to explore molecular mechanisms in cotton species. A total of 1502 GELP genes were identified in 10 cotton species. Segmental duplication and differences in evolutionary rates are the leading causes of the increase in the number and diversity of GELP genes during evolution for ecological adaptation. Structural analysis revealed that the GELP family has high structural diversity. Moreover, molecular simulation studies have demonstrated significant differences in the properties of the binding pockets among cotton GELPs. In the process of adapting to the environment, GELPs not only have segmental duplication but also have different evolutionary rates, resulting in gene diversity. This diversity leads to significant differences in the 3D structure and binding pocket properties and, finally, to functional diversity. These findings provide a reference for further functional analyses of plant GELPs.
Keywords: GDSL-type esterases/lipases; Gossypium species; binding pocket; functional diversity; molecular mechanism; molecular simulation.
Copyright © 2023 Wang, Zhao, Qu, Yang and Huang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







Similar articles
-
Identification, evolution, and expression of GDSL-type Esterase/Lipase (GELP) gene family in three cotton species: a bioinformatic analysis.BMC Genomics. 2023 Dec 21;24(1):795. doi: 10.1186/s12864-023-09717-3. BMC Genomics. 2023. PMID: 38129780 Free PMC article.
-
Expansion and evolutionary patterns of GDSL-type esterases/lipases in Rosaceae genomes.Funct Integr Genomics. 2018 Nov;18(6):673-684. doi: 10.1007/s10142-018-0620-1. Epub 2018 Jun 13. Funct Integr Genomics. 2018. PMID: 29948460
-
Key Amino Acids for Transferase Activity of GDSL Lipases.Int J Mol Sci. 2022 Dec 1;23(23):15141. doi: 10.3390/ijms232315141. Int J Mol Sci. 2022. PMID: 36499468 Free PMC article.
-
Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development.Plants (Basel). 2022 Feb 9;11(4):468. doi: 10.3390/plants11040468. Plants (Basel). 2022. PMID: 35214802 Free PMC article. Review.
-
Genome-Wide Classification and Phylogenetic Analyses of the GDSL-Type Esterase/Lipase (GELP) Family in Flowering Plants.Int J Mol Sci. 2022 Oct 11;23(20):12114. doi: 10.3390/ijms232012114. Int J Mol Sci. 2022. PMID: 36292971 Free PMC article. Review.
Cited by
-
Comparative omics-based characterization, phylogeny and melatonin-mediated expression analyses of GDSL genes in pitaya (Selenicereus undatus L.) against multifactorial abiotic stresses.Physiol Mol Biol Plants. 2024 Sep;30(9):1493-1515. doi: 10.1007/s12298-024-01506-w. Epub 2024 Sep 5. Physiol Mol Biol Plants. 2024. PMID: 39310703 Free PMC article.
-
Towards carotenoid biofortification in wheat: identification of XAT-7A1, a multicopy tandem gene responsible for carotenoid esterification in durum wheat.BMC Plant Biol. 2023 Sep 6;23(1):412. doi: 10.1186/s12870-023-04431-4. BMC Plant Biol. 2023. PMID: 37674126 Free PMC article.
-
Transcriptomic and Metabolomic Profiling of Root Tissue in Drought-Tolerant and Drought-Susceptible Wheat Genotypes in Response to Water Stress.Int J Mol Sci. 2024 Sep 27;25(19):10430. doi: 10.3390/ijms251910430. Int J Mol Sci. 2024. PMID: 39408761 Free PMC article.
-
Microenzymes: Is There Anybody Out There?Protein J. 2024 Jun;43(3):393-404. doi: 10.1007/s10930-024-10193-1. Epub 2024 Mar 20. Protein J. 2024. PMID: 38507106 Review.
-
Identification, evolution, and expression of GDSL-type Esterase/Lipase (GELP) gene family in three cotton species: a bioinformatic analysis.BMC Genomics. 2023 Dec 21;24(1):795. doi: 10.1186/s12864-023-09717-3. BMC Genomics. 2023. PMID: 38129780 Free PMC article.
References
-
- An J., Ze P., Li H. (2017). GDSL lipase increases the resistance against Verticillium dahilae in Gossypium hirsutum . J. Shihezi Univ. Natural Science. 35, 420–424. doi: 10.13880/j.cnki.65-1174/n.2017.04.005 - DOI
LinkOut - more resources
Full Text Sources