

Hexavalent chromium-induced epigenetic instability and transposon activation lead to phenotypic variations and tumors in Drosophila
- PMID: 36743586
- PMCID: PMC9892686
- DOI: 10.1093/eep/dvac030
Hexavalent chromium-induced epigenetic instability and transposon activation lead to phenotypic variations and tumors in Drosophila
Abstract
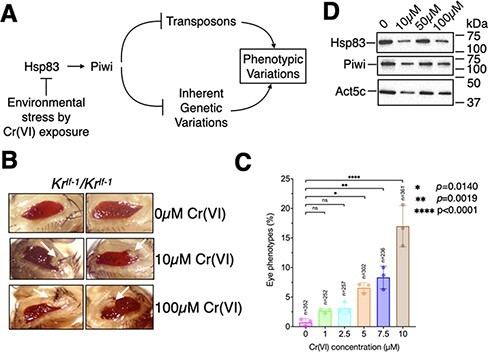
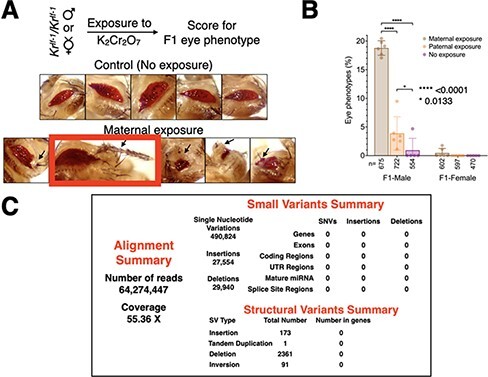
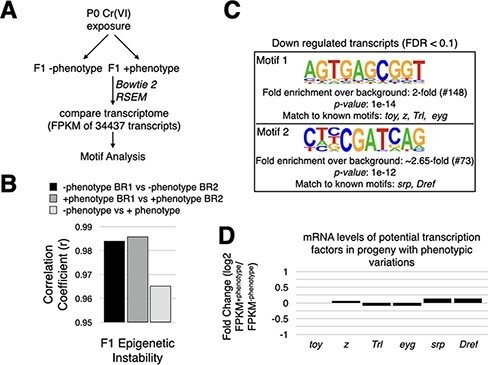
Developmental robustness represents the ability of an organism to resist phenotypic variations despite environmental insults and inherent genetic variations. Derailment of developmental robustness leads to phenotypic variations that can get fixed in a population for many generations. Environmental pollution is a significant worldwide problem with detrimental consequences of human development. Understanding the genetic basis for how pollutants affect human development is critical for developing interventional therapies. Here, we report that environmental stress induced by hexavalent chromium, Cr(VI), a potent industrial pollutant, compromises developmental robustness, leading to phenotypic variations in the progeny. These phenotypic variations arise due to epigenetic instability and transposon activation in the somatic tissues of the progeny rather than novel genetic mutations and can be reduced by increasing the dosage of Piwi - a Piwi-interacting RNA-binding protein, in the ovary of the exposed mother. Significantly, the derailment of developmental robustness by Cr(VI) exposure leads to tumors in the progeny, and the predisposition to develop tumors is fixed in the population for at least three generations. Thus, we show for the first time that environmental pollution can derail developmental robustness and predispose the progeny of the exposed population to develop phenotypic variations and tumors.
Keywords: canalization; developmental robustness; epigenetic inheritance; epigenetic instability; hexavalent chromium; tumor.
© The Author(s) 2023. Published by Oxford University Press.
Conflict of interest statement
None declared.
Figures



Similar articles
-
Drosophila Piwi functions in Hsp90-mediated suppression of phenotypic variation.Nat Genet. 2011 Feb;43(2):153-8. doi: 10.1038/ng.743. Epub 2010 Dec 26. Nat Genet. 2011. PMID: 21186352 Free PMC article.
-
Environmental Presence of Hexavalent but Not Trivalent Chromium Causes Neurotoxicity in Exposed Drosophila melanogaster.Mol Neurobiol. 2017 Jul;54(5):3368-3387. doi: 10.1007/s12035-016-9909-z. Epub 2016 May 11. Mol Neurobiol. 2017. PMID: 27167131
-
Epigenetic downregulation of O6-methylguanine-DNA methyltransferase contributes to chronic hexavalent chromium exposure-caused genotoxic effect and cell transformation.Environ Pollut. 2024 Jan 15;341:122978. doi: 10.1016/j.envpol.2023.122978. Epub 2023 Nov 21. Environ Pollut. 2024. PMID: 37995958 Free PMC article.
-
Untangling the web: the diverse functions of the PIWI/piRNA pathway.Mol Reprod Dev. 2013 Aug;80(8):632-64. doi: 10.1002/mrd.22195. Epub 2013 Jun 27. Mol Reprod Dev. 2013. PMID: 23712694 Free PMC article. Review.
-
Health hazards of hexavalent chromium (Cr (VI)) and its microbial reduction.Bioengineered. 2022 Mar;13(3):4923-4938. doi: 10.1080/21655979.2022.2037273. Bioengineered. 2022. PMID: 35164635 Free PMC article. Review.
References
-
- Waddington CH. Canalization of development and genetic assimilation of acquired characters. Nature 1959;183:1654–5. - PubMed
-
- Rutherford SL, Lindquist S. Hsp90 as a capacitor for morphological evolution. Nature 1998;396:336–42. - PubMed
-
- Sollars V, Lu X, Xiao L et al. Evidence for an epigenetic mechanism by which Hsp90 acts as a capacitor for morphological evolution. Nat Genet 2003;33:70–4. - PubMed
-
- Hummel B, Hansen EC, Yoveva A et al. The evolutionary capacitor HSP90 buffers the regulatory effects of mammalian endogenous retroviruses. Nat Struct Mol Biol 2017;24:234–42. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
