

Repeated Emergence of Variant TetR Family Regulator, FarR, and Increased Resistance to Antimicrobial Unsaturated Fatty Acid among Clonal Complex 5 Methicillin-Resistant Staphylococcus aureus
- PMID: 36744906
- PMCID: PMC10019231
- DOI: 10.1128/aac.00749-22
Repeated Emergence of Variant TetR Family Regulator, FarR, and Increased Resistance to Antimicrobial Unsaturated Fatty Acid among Clonal Complex 5 Methicillin-Resistant Staphylococcus aureus
Abstract
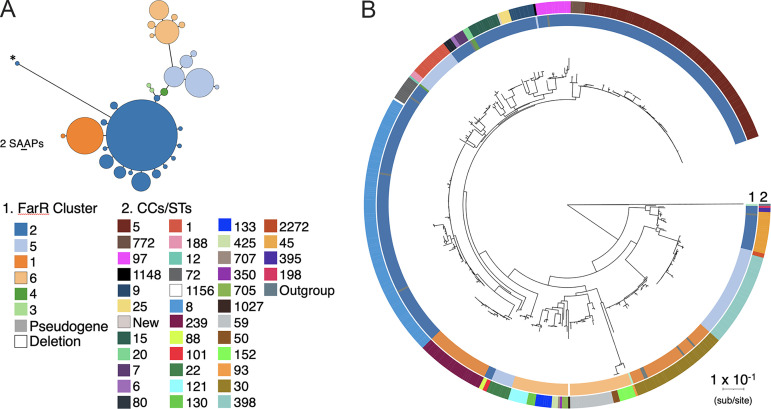
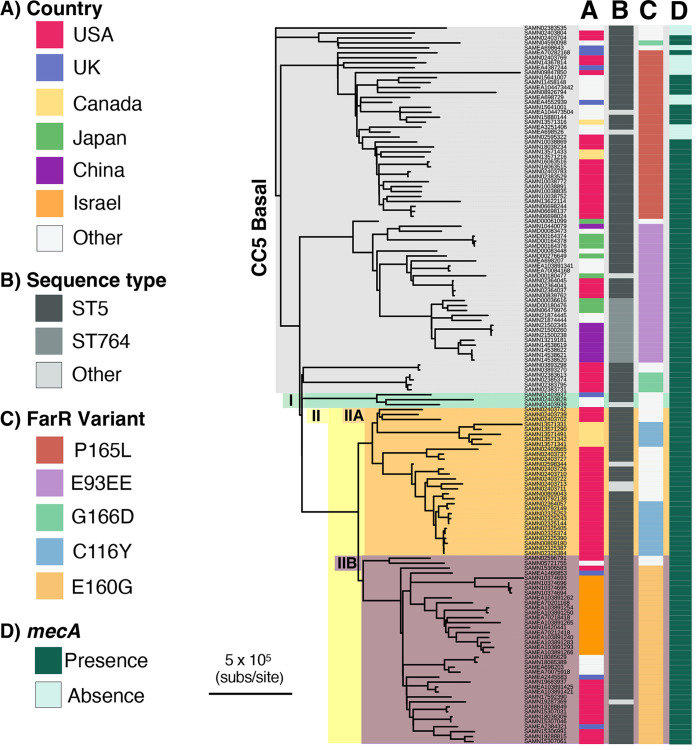
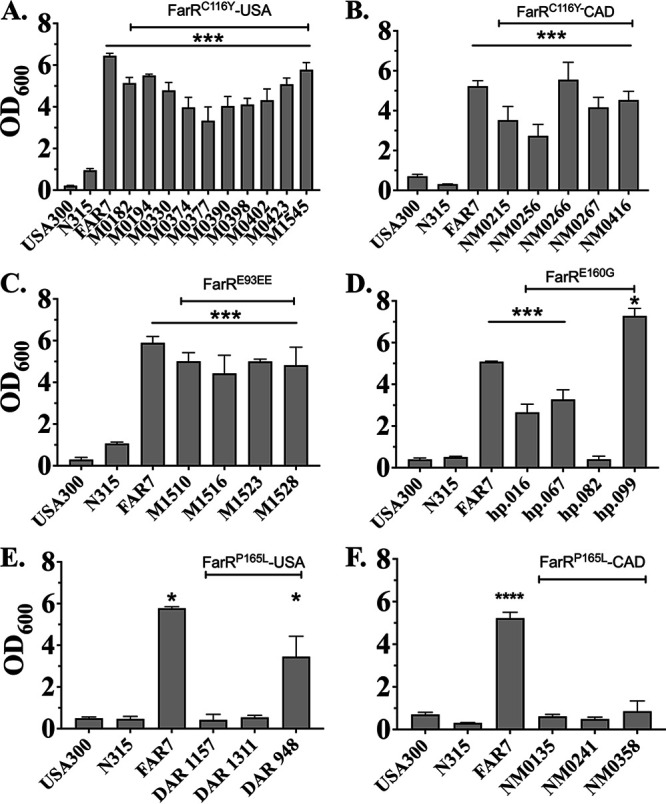
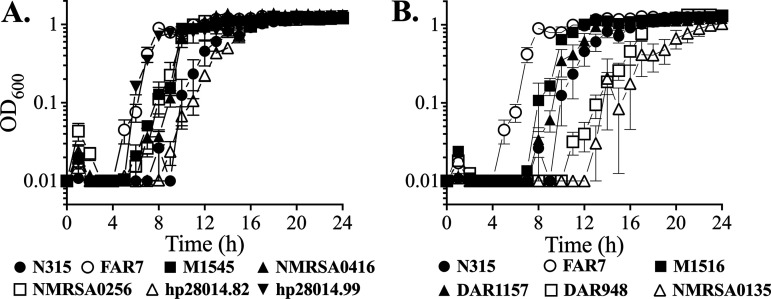
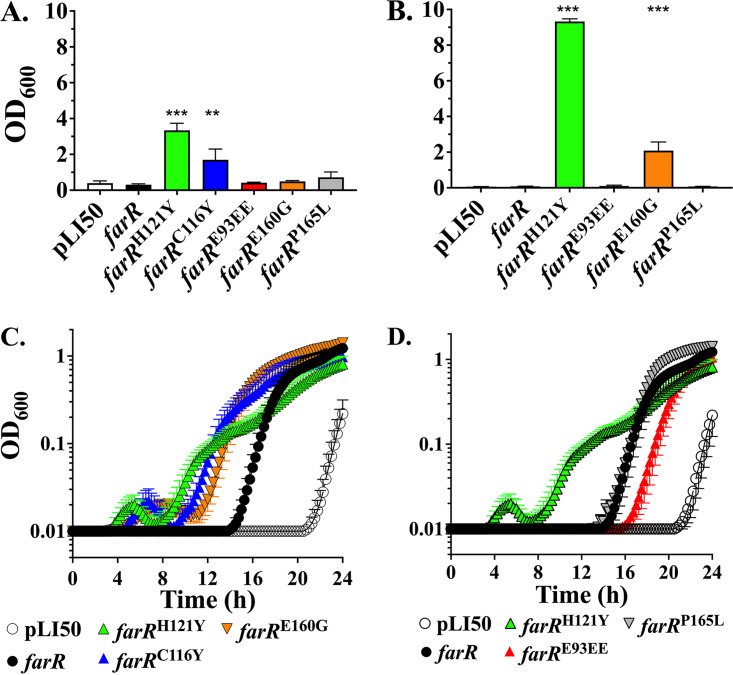
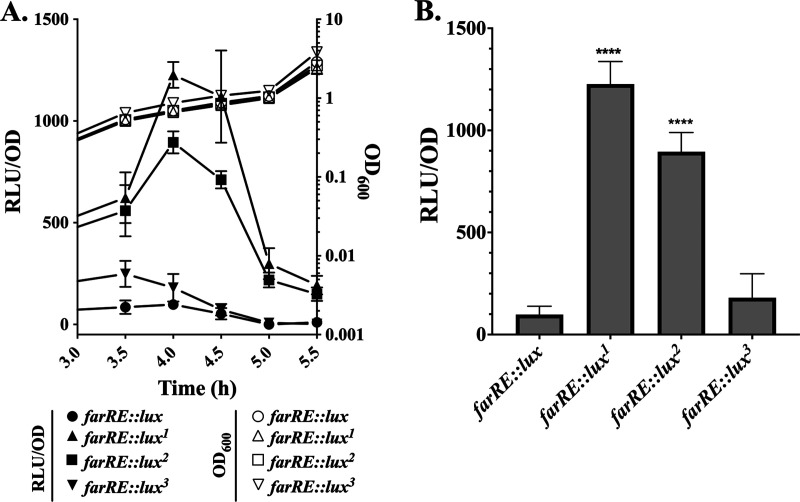
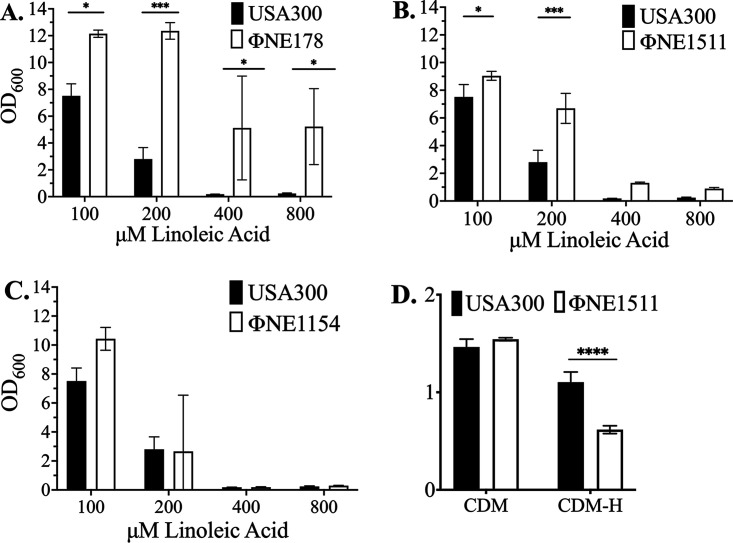
Resistance-nodulation-division (RND) superfamily efflux pumps promote antibiotic resistance in Gram-negative pathogens, but their role in Gram-positive pathogens, including methicillin-resistant Staphylococcus aureus (MRSA) is undocumented. However, recent in vitro selections for resistance of S. aureus to an antimicrobial fatty acid, linoleic acid, and an antibiotic, rhodomyrtone, identified H121Y and C116R substitution variants, respectively, in a TetR family regulator, FarR, promoting increased expression of the RND pump FarE. Hypothesizing that in vivo selection pressures have also promoted the emergence of FarR variants, we searched available genome data and found that strains with FarRH121Y from human and bovine hosts have emerged sporadically in clonal complexes (CCs) CC1, CC30, CC8, CC22, and CC97, whereas multiple FarR variants have occurred within CC5 hospital-associated (HA)-MRSA. Of these, FarRE160G and FarRE93EE were exclusive to CC5, while FarRC116Y, FarRP165L, and FarRG166D also occurred in nonrelated CCs, primarily from bovine hosts. Within CC5, FarRC116Y and FarRG166D strains were polyphyletic, each exhibiting two emergence events. FarRC116Y and FarRE160G were individually sufficient to confer increased expression of FarE and enhanced resistance to linoleic acid (LA). Isolates with FarRE93EE were most closely related to S. aureus N315 MRSA and exhibited increased resistance independently of FarRE93EE. Accumulation of pseudogenes and additional polymorphisms in FarRE93EE strains contributed to a multiresistance phenotype which included fosfomycin and fusidic acid resistance in addition to increased linoleic acid resistance. These findings underscore the remarkable adaptive capacity of CC5 MRSA, which includes the polyphyletic USA100 lineage of HA-MRSA that is endemic in the Western hemisphere and known for the acquisition of multiple resistance phenotypes.
Keywords: MRSA; Staphylococcus aureus; TetR family regulator; efflux pumps; mechanisms of resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
