

Paradoxical electro-olfactogram responses in TMEM16B knock-out mice
- PMID: 36744918
- PMCID: PMC9951260
- DOI: 10.1093/chemse/bjad003
Paradoxical electro-olfactogram responses in TMEM16B knock-out mice
Abstract
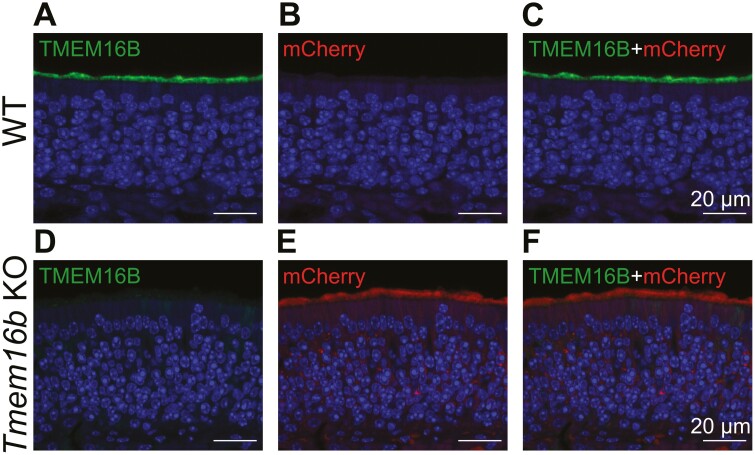
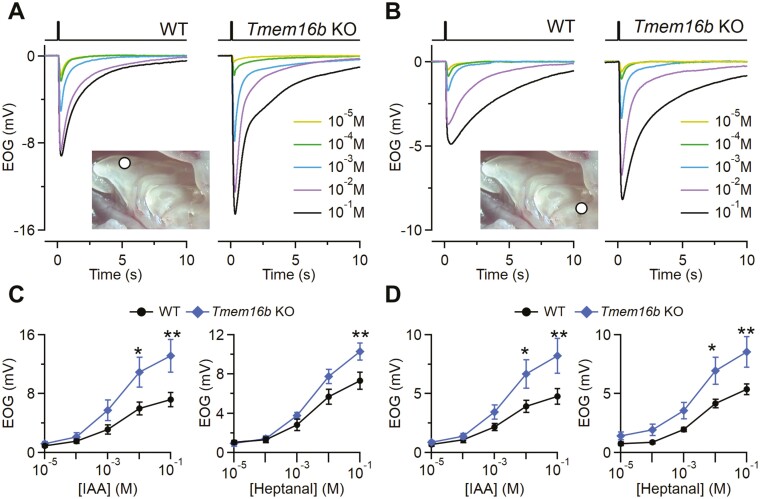
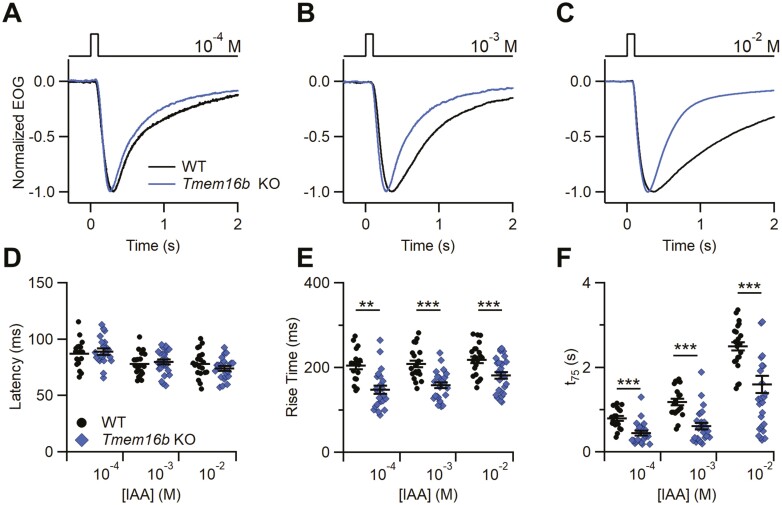
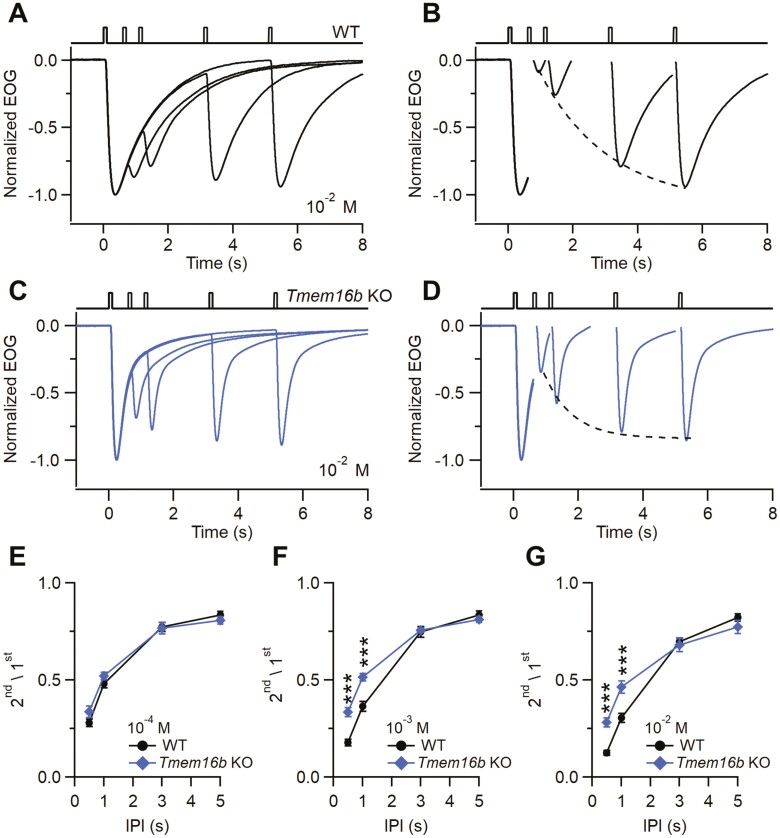
The Ca2+-activated Cl¯ channel TMEM16B carries up to 90% of the transduction current evoked by odorant stimulation in olfactory sensory neurons and control the number of action potential firing and therefore the length of the train of action potentials. A loss of function approach revealed that TMEM16B is required for olfactory-driven behaviors such as tracking unfamiliar odors. Here, we used the electro-olfactogram (EOG) technique to investigate the contribution of TMEM16B to odorant transduction in the whole olfactory epithelium. Surprisingly, we found that EOG responses from Tmem16b knock out mice have a bigger amplitude compared to those of wild type. Moreover, the kinetics of EOG responses is faster in absence of TMEM16B, while the ability to adapt to repeated stimulation is altered in knock out mice. The larger EOG responses in Tmem16b knock out may be the results of the removal of the clamping and/or shunting action of the Ca2+-activated Cl¯ currents leading to the paradox of having smaller transduction current but larger generator potential.
Keywords: ANO2; Ca2+-activated Cl¯ currents; olfactory sensory neurons; olfactory transduction.
© The Author(s) 2023. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Belluscio L, Gold GH, Nemes A, Axel R.. Mice deficient in G(olf) are anosmic. Neuron. 1998;20:69–81. - PubMed
-
- Billig GM, Pál B, Fidzinski P, Jentsch TJ.. Ca2+-activated Cl− currents are dispensable for olfaction. Nat Neurosci. 2011;14:763–769. - PubMed
-
- Boccaccio A, Menini A.. Temporal development of cyclic nucleotide-gated and Ca2+ -activated Cl- currents in isolated mouse olfactory sensory neurons. J Neurophysiol. 2007;98:153–160. - PubMed
-
- Boccaccio, A, Menini, A., Pifferi, S.. The cyclic AMP signaling pathway in the rodent main olfactory system. Cell Tissue Res. 2021;383:429–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
