

Defective excitation-contraction coupling and mitochondrial respiration precede mitochondrial Ca2+ accumulation in spinobulbar muscular atrophy skeletal muscle
- PMID: 36746942
- PMCID: PMC9902403
- DOI: 10.1038/s41467-023-36185-w
Defective excitation-contraction coupling and mitochondrial respiration precede mitochondrial Ca2+ accumulation in spinobulbar muscular atrophy skeletal muscle
Abstract
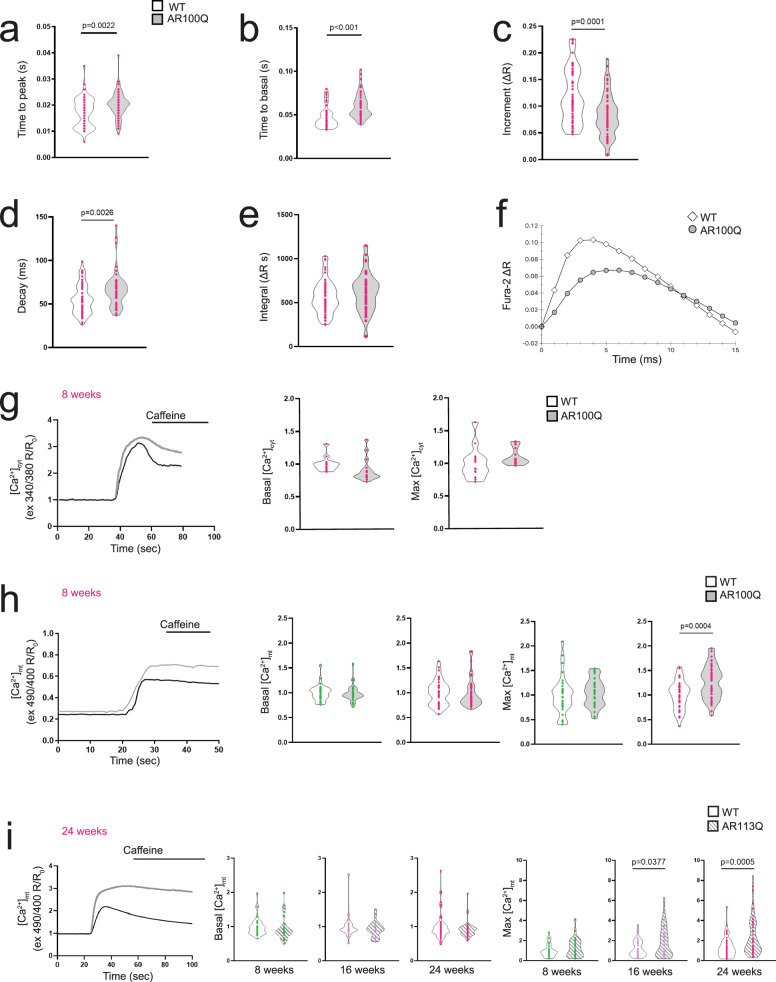
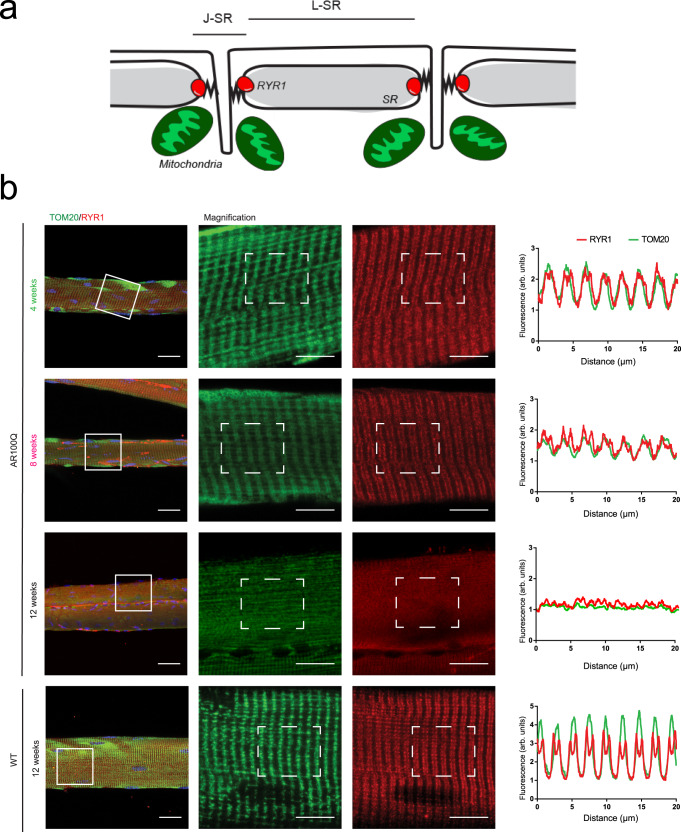
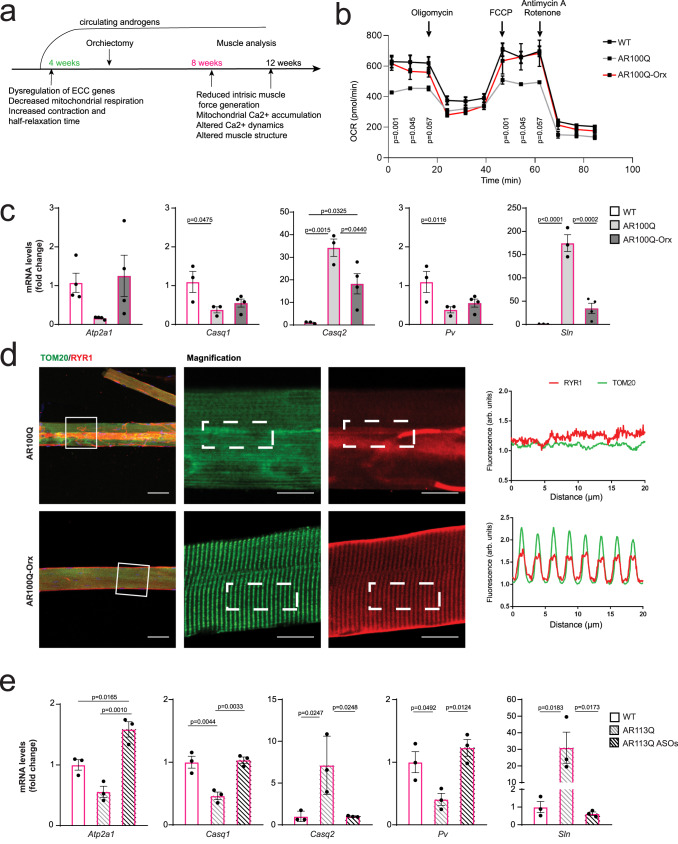
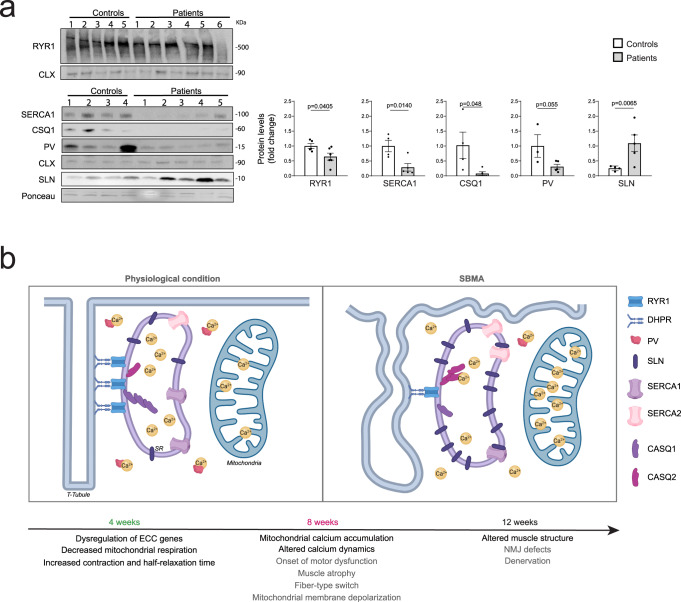
Polyglutamine expansion in the androgen receptor (AR) causes spinobulbar muscular atrophy (SBMA). Skeletal muscle is a primary site of toxicity; however, the current understanding of the early pathological processes that occur and how they unfold during disease progression remains limited. Using transgenic and knock-in mice and patient-derived muscle biopsies, we show that SBMA mice in the presymptomatic stage develop a respiratory defect matching defective expression of genes involved in excitation-contraction coupling (ECC), altered contraction dynamics, and increased fatigue. These processes are followed by stimulus-dependent accumulation of calcium into mitochondria and structural disorganization of the muscle triads. Deregulation of expression of ECC genes is concomitant with sexual maturity and androgen raise in the serum. Consistent with the androgen-dependent nature of these alterations, surgical castration and AR silencing alleviate the early and late pathological processes. These observations show that ECC deregulation and defective mitochondrial respiration are early but reversible events followed by altered muscle force, calcium dyshomeostasis, and dismantling of triad structure.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
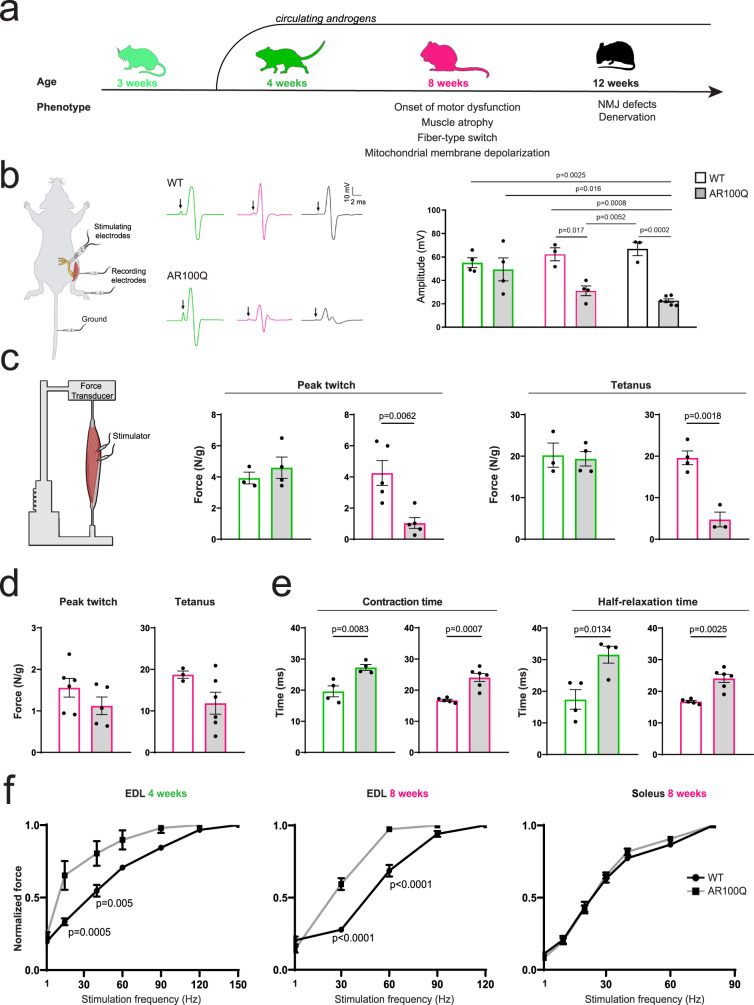
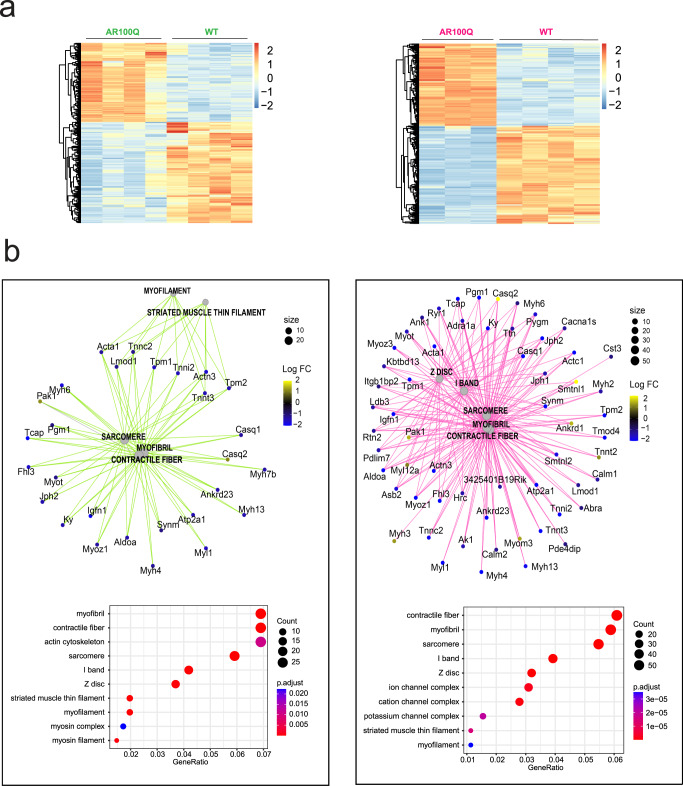
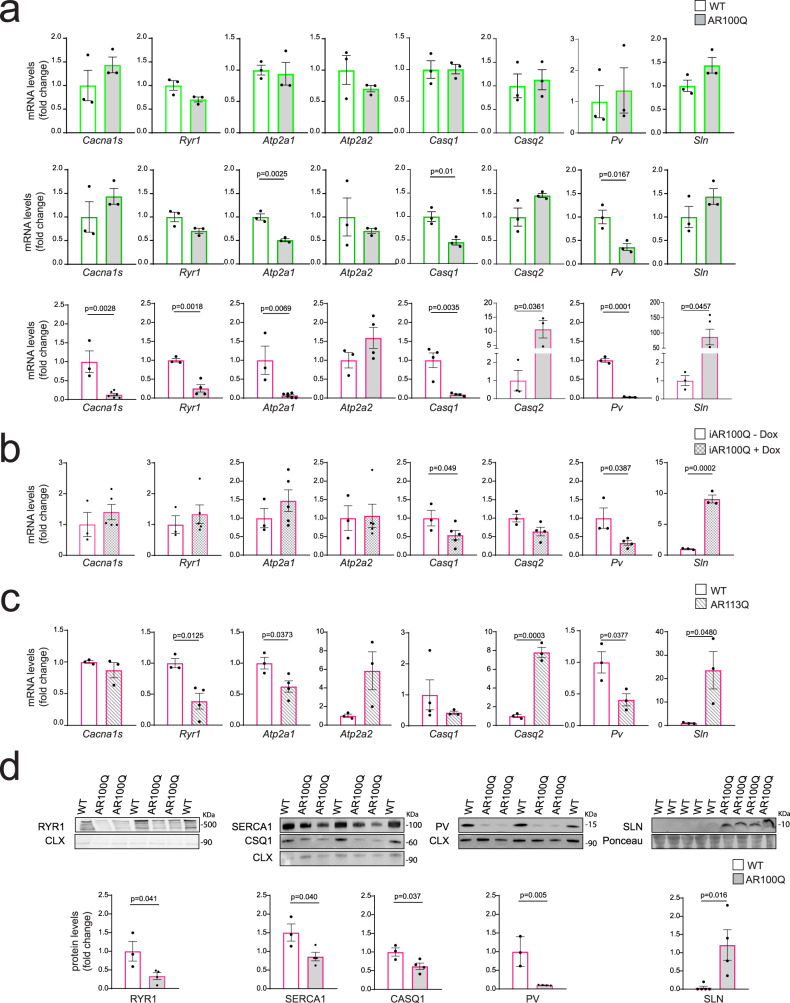
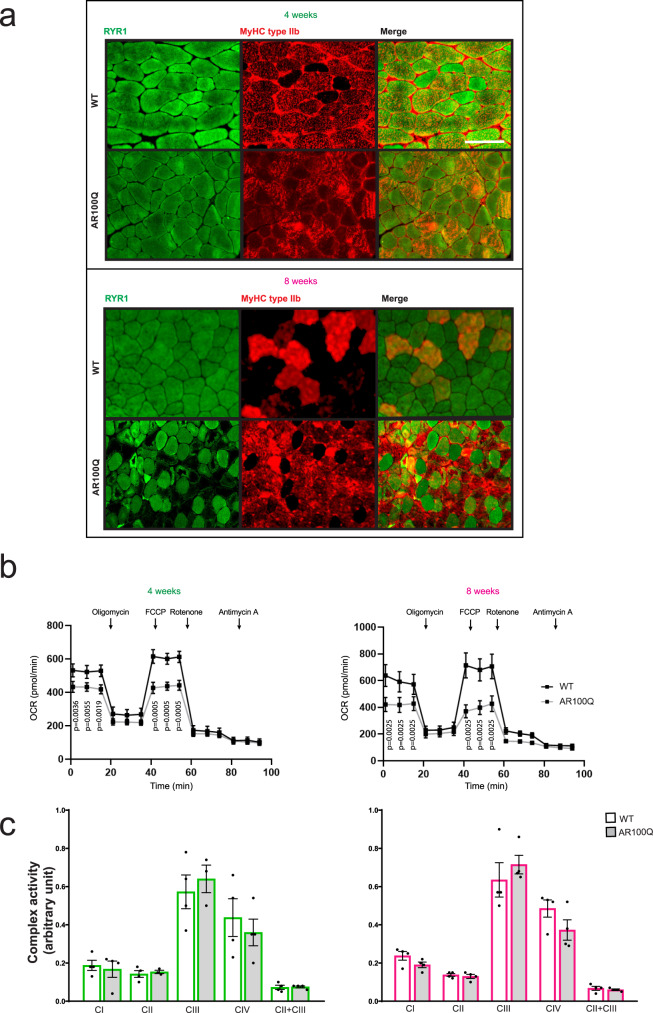
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
