Rapid diversification underlying the global dominance of a cosmopolitan phytoplankton
- PMID: 36747097
- PMCID: PMC10030636
- DOI: 10.1038/s41396-023-01365-5
Rapid diversification underlying the global dominance of a cosmopolitan phytoplankton
Abstract
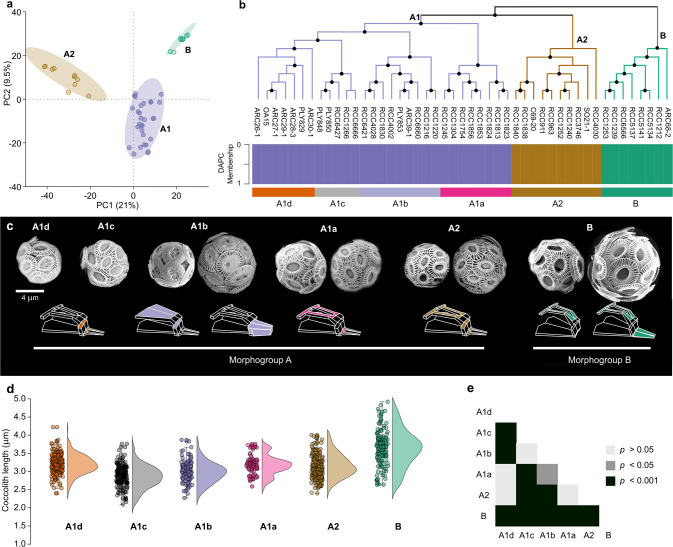
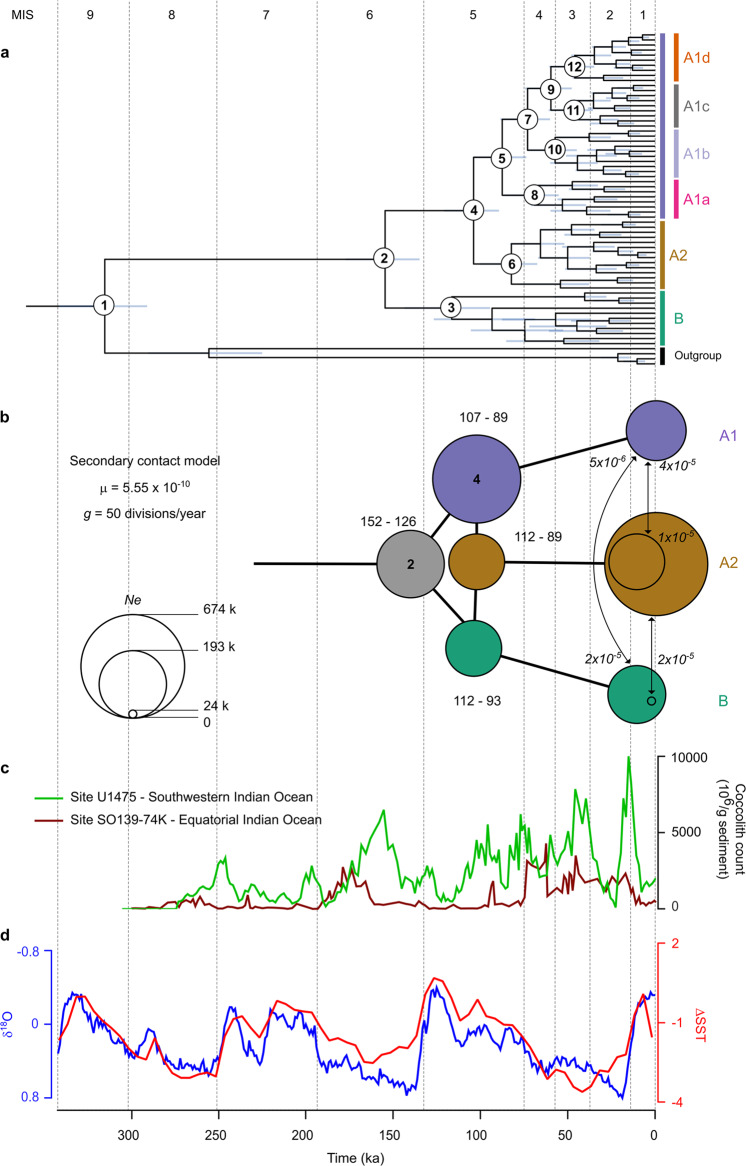
Marine phytoplankton play important roles in the global ecosystem, with a limited number of cosmopolitan keystone species driving their biomass. Recent studies have revealed that many of these phytoplankton are complexes composed of sibling species, but little is known about the evolutionary processes underlying their formation. Gephyrocapsa huxleyi, a widely distributed and abundant unicellular marine planktonic algae, produces calcified scales (coccoliths), thereby significantly affects global biogeochemical cycles via sequestration of inorganic carbon. This species is composed of morphotypes defined by differing degrees of coccolith calcification, the evolutionary ecology of which remains unclear. Here, we report an integrated morphological, ecological and genomic survey across globally distributed G. huxleyi strains to reconstruct evolutionary relationships between morphotypes in relation to their habitats. While G. huxleyi has been considered a single cosmopolitan species, our analyses demonstrate that it has evolved to comprise at least three distinct species, which led us to formally revise the taxonomy of the G. huxleyi complex. Moreover, the first speciation event occurred before the onset of the last interglacial period (~140 ka), while the second followed during this interglacial. Then, further rapid diversifications occurred during the most recent ice-sheet expansion of the last glacial period and established morphotypes as dominant populations across environmental clines. These results suggest that glacial-cycle dynamics contributed to the isolation of ocean basins and the segregations of oceans fronts as extrinsic drivers of micro-evolutionary radiations in extant marine phytoplankton.
© 2023. Crown.
Conflict of interest statement
The authors declare no competing interests.
Figures