

This is a preprint.
Targeting Immune-Fibroblast Crosstalk in Myocardial Infarction and Cardiac Fibrosis
- PMID: 36747878
- PMCID: PMC9900986
- DOI: 10.21203/rs.3.rs-2402606/v1
Targeting Immune-Fibroblast Crosstalk in Myocardial Infarction and Cardiac Fibrosis
Update in
-
Targeting immune-fibroblast cell communication in heart failure.Nature. 2024 Nov;635(8038):423-433. doi: 10.1038/s41586-024-08008-5. Epub 2024 Oct 23. Nature. 2024. PMID: 39443792 Free PMC article.
Abstract
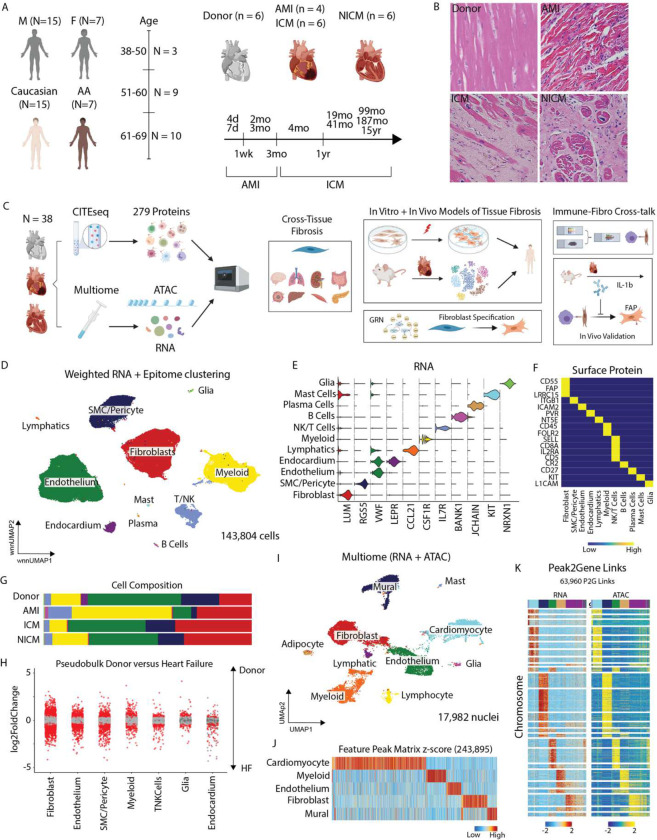
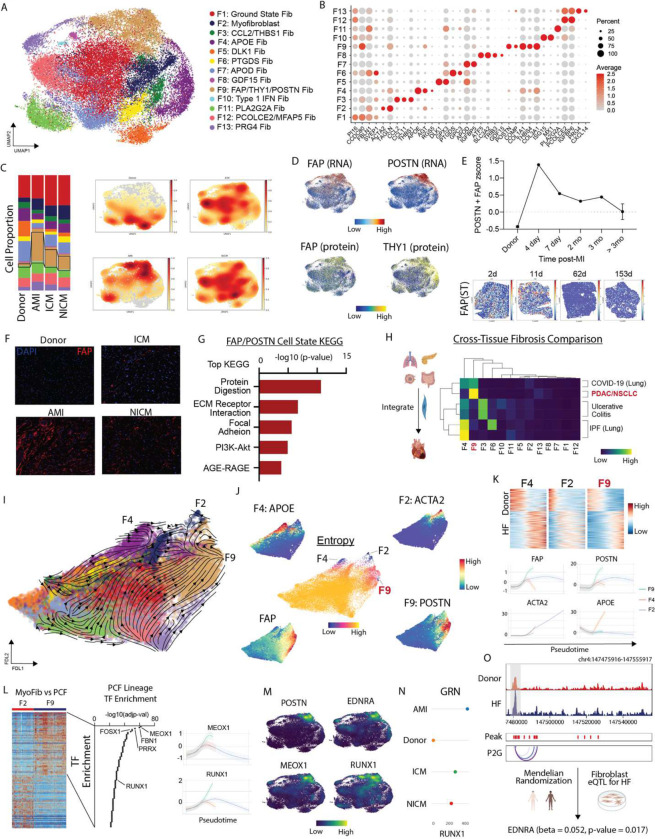
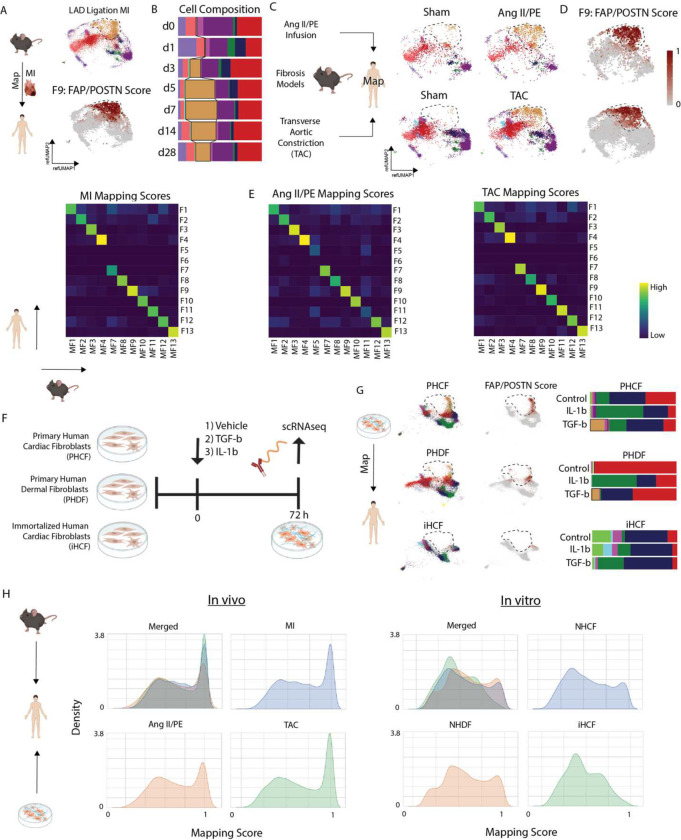
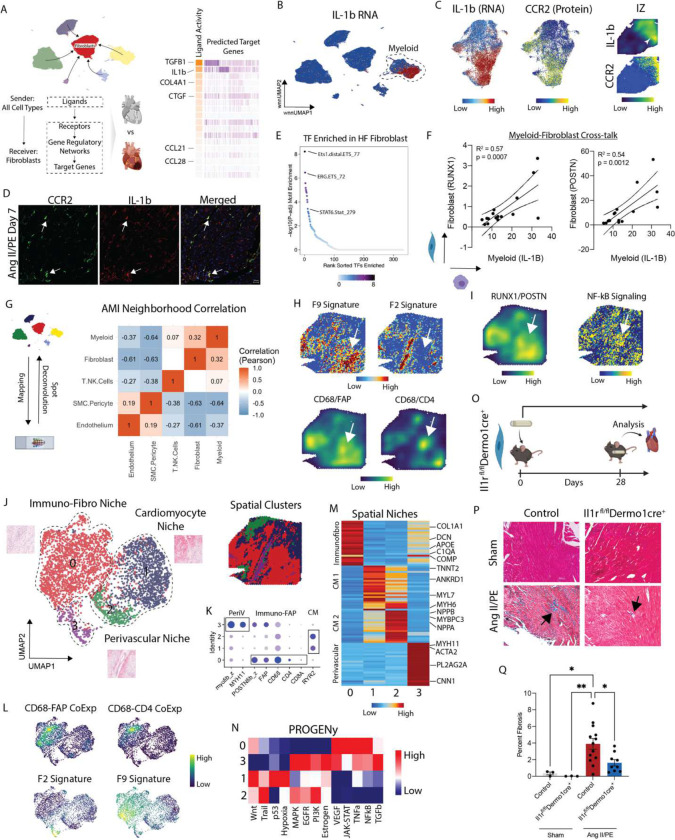
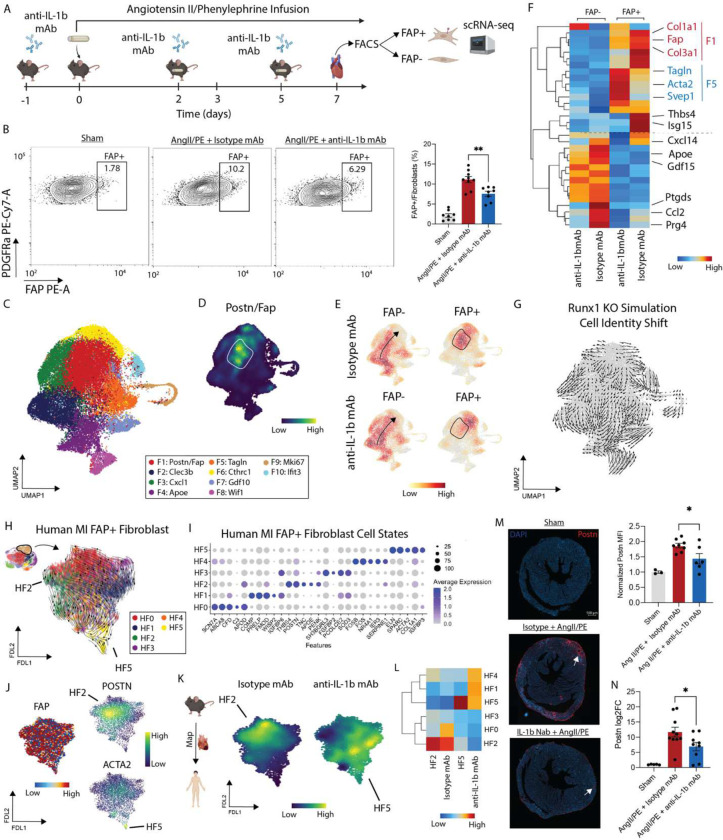
Inflammation and tissue fibrosis co-exist and are causally linked to organ dysfunction. However, the molecular mechanisms driving immune-fibroblast crosstalk in human cardiac disease remains unexplored and there are currently no therapeutics to target fibrosis. Here, we performed multi-omic single-cell gene expression, epitope mapping, and chromatin accessibility profiling in 38 donors, acutely infarcted, and chronically failing human hearts. We identified a disease-associated fibroblast trajectory marked by cell surface expression of fibroblast activator protein (FAP), which diverged into distinct myofibroblasts and pro-fibrotic fibroblast populations, the latter resembling matrifibrocytes. Pro-fibrotic fibroblasts were transcriptionally similar to cancer associated fibroblasts and expressed high levels of collagens and periostin (POSTN), thymocyte differentiation antigen 1 (THY-1), and endothelin receptor A (EDNRA) predicted to be driven by a RUNX1 gene regulatory network. We assessed the applicability of experimental systems to model tissue fibrosis and demonstrated that 3 different in vivo mouse models of cardiac injury were superior compared to cultured human heart and dermal fibroblasts in recapitulating the human disease phenotype. Ligand-receptor analysis and spatial transcriptomics predicted that interactions between C-C chemokine receptor type 2 (CCR2) macrophages and fibroblasts mediated by interleukin 1 beta (IL-1β) signaling drove the emergence of pro-fibrotic fibroblasts within spatially defined niches. This concept was validated through in silico transcription factor perturbation and in vivo inhibition of IL-1β signaling in fibroblasts where we observed reduced pro-fibrotic fibroblasts, preferential differentiation of fibroblasts towards myofibroblasts, and reduced cardiac fibrosis. Herein, we show a subset of macrophages signal to fibroblasts via IL-1β and rewire their gene regulatory network and differentiation trajectory towards a pro-fibrotic fibroblast phenotype. These findings highlight the broader therapeutic potential of targeting inflammation to treat tissue fibrosis and restore organ function.
Keywords: C-C chemokine receptor 2; fibroblast activator protein; fibrosis; heart failure; interleukin 1 beta; macrophages.
Conflict of interest statement
Competing Interests XL, TY, SS, AF, MF, C-ML and BA are or were employed by Amgen.
Figures
References
-
- Gutstein D. E. & Fuster V. Pathophysiology and clinical significance of atherosclerotic plaque rupture. Cardiovasc. Res. 41, 323–333 (1999). - PubMed
-
- Thygesen K. et al. Third universal definition of myocardial infarction. Circulation 126, 2020–2035 (2012). - PubMed
-
- Gómez A. M., Guatimosim S., Dilly K. W., Vassort G. & Lederer W. j. Heart Failure After Myocardial Infarction. Circulation 104, 688–693 (2001). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
