

Interspecies Horizontal Transfer and Specific Integration of the Mosquitocidal Toxin-Encoding Plasmid pTAND672-2 from Bacillus thuringiensis subsp. israelensis to Lysinibacillus sphaericus
- PMID: 36749061
- PMCID: PMC9973010
- DOI: 10.1128/aem.01652-22
Interspecies Horizontal Transfer and Specific Integration of the Mosquitocidal Toxin-Encoding Plasmid pTAND672-2 from Bacillus thuringiensis subsp. israelensis to Lysinibacillus sphaericus
Abstract
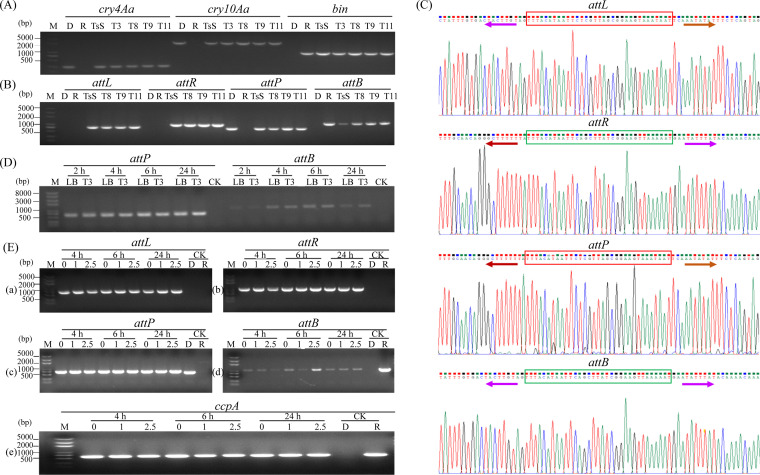
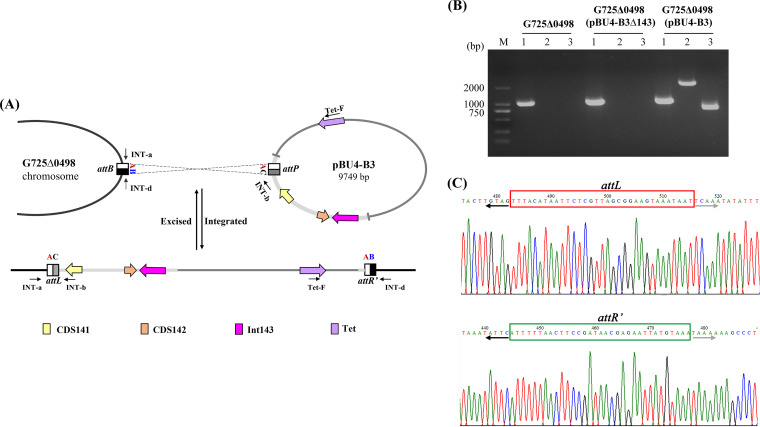
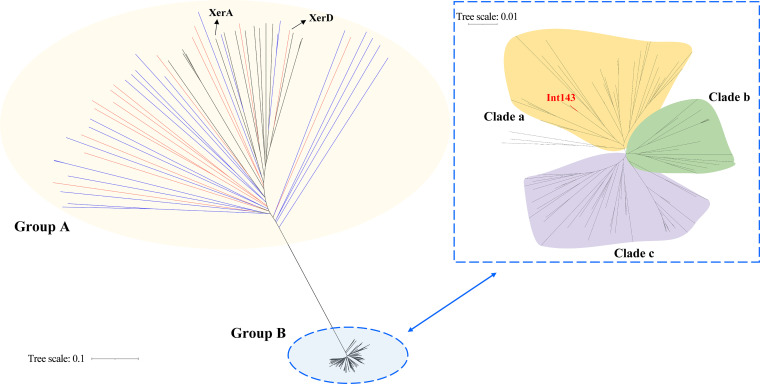
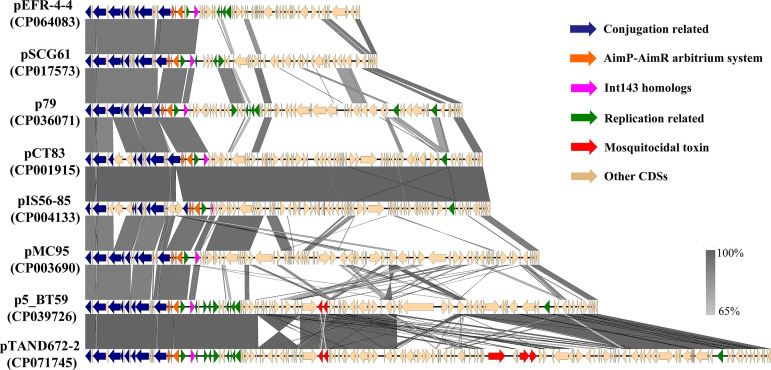
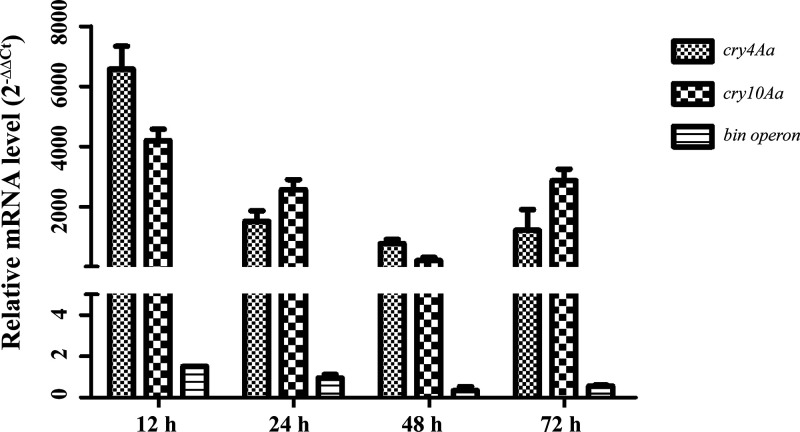
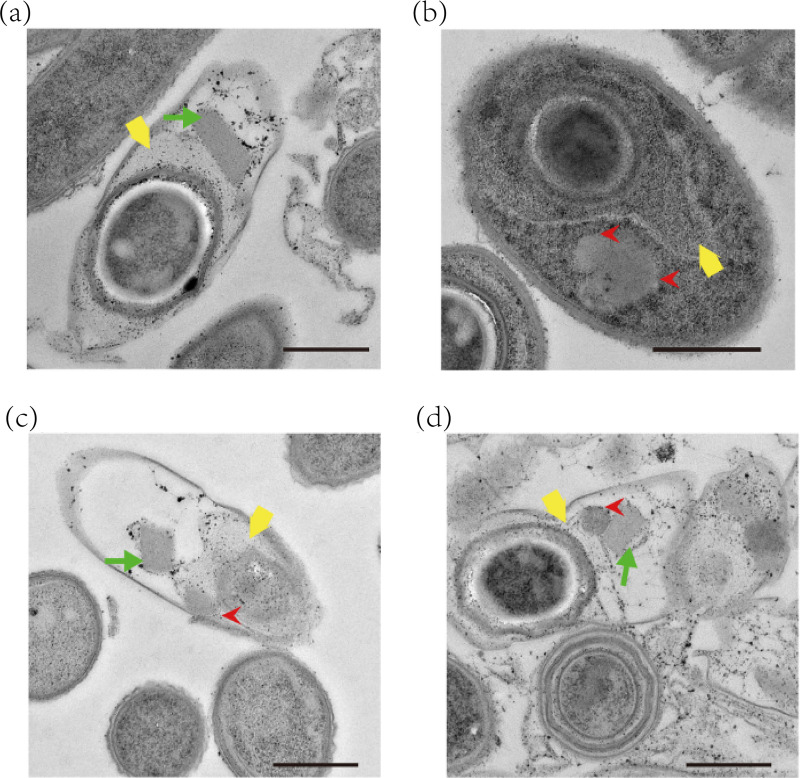
pTAND672-2, a 144-kb resident plasmid of Bacillus thuringiensis serovar israelensis strain TAND672, was sequenced and characterized. This extrachromosomal element carries mosquitocidal toxin-, conjugation-, and recombinase-encoding genes, together with a putative arbitrium system, a genetic module recently discovered in temperate phages controlling lysogeny-lysis transition and in mobile genetic elements (MGEs) where its function remains clarified. Using conjugation experiments, pTAND672-2 is shown to be a novel integrative and conjugative element (ICE), which can horizontally transfer from B. thuringiensis serovar israelensis to Lysinibacillus sphaericus, another mosquitocidal bacterium, where it integrates into the chromosome. Its integration and circularization are reversible and involve a single-cross recombination between 33-bp specific sites, attB in the chromosome of L. sphaericus and attP in pTAND672-2. CDS143, coding for the putative tyrosine integrase Int143 distantly related to site-specific tyrosine Xer recombinases and phage integrases, can mediate the integration of pTAND672-2 to attB. The B. thuringiensis mosquito-killing genes carried by pTAND672-2 are efficiently transcribed and expressed in L. sphaericus, displaying a slight increased toxicity in this bacterium against Aedes albopictus larvae. The occurrence of pTAND672-2-like plasmids within the Bacillus cereus group was also explored and indicated that they all share a similar genetic backbone with diverse plasmid sizes, ranging from 58 to 225 kb. Interestingly, among them, the pEFR-4-4 plasmid of Bacillus paranthracis EFR-4 and p5 of B. thuringiensis BT-59 also display conjugative capability; moreover, like pTAND672-2 displays a chimeric structure between the pCH_133-e- and pBtoxis-like plasmids, pBTHD789-3 also appears to be mosaic of two plasmids. IMPORTANCE Horizontal transfer of mobile genetic elements carrying mosquitocidal toxin genes may play a driving role in the diversity of mosquitocidal bacteria. Here, the 144-kb mosquitocidal toxin-encoding plasmid pTAND672-2 is the first verified integrative and conjugative element (ICE) identified in Bacillus thuringiensis serovar israelensis. The key tyrosine integrase Int143, involved in the specific integration, is distantly related to other tyrosine recombinases. The study also reports the occurrence and potential interspecies transmission of pTAND672-2-like plasmids with varied sizes in B. thuringiensis, Bacillus paranthracis, and Bacillus wiedmannii isolates belonging to the Bacillus cereus group. This study is important for further understanding the evolution and ecology of mosquitocidal bacteria, as well as for providing new direction for the genetic engineering of biopesticides in the control of disease-transmitting mosquitoes.
Keywords: Bacillus thuringiensis serovar israelensis; Lysinibacillus sphaericus; conjugation; integrative and conjugative element; mobile genetic elements; mosquitocidal toxin; tyrosine recombinase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Conjugal transfer of a toxin-coding megaplasmid from Bacillus thuringiensis subsp. israelensis to mosquitocidal strains of Bacillus sphaericus.Appl Environ Microbiol. 2006 Mar;72(3):1766-70. doi: 10.1128/AEM.72.3.1766-1770.2006. Appl Environ Microbiol. 2006. PMID: 16517620 Free PMC article.
-
Molecular cloning of the 130-kilodalton mosquitocidal delta-endotoxin gene of Bacillus thuringiensis subsp. israelensis in Bacillus sphaericus.Appl Environ Microbiol. 1990 Jun;56(6):1710-6. doi: 10.1128/aem.56.6.1710-1716.1990. Appl Environ Microbiol. 1990. PMID: 2200339 Free PMC article.
-
Improvement of Bacillus sphaericus toxicity against dipteran larvae by integration, via homologous recombination, of the Cry11A toxin gene from Bacillus thuringiensis subsp. israelensis.Appl Environ Microbiol. 1997 Nov;63(11):4413-20. doi: 10.1128/aem.63.11.4413-4420.1997. Appl Environ Microbiol. 1997. PMID: 9361428 Free PMC article.
-
Microbial control of mosquitoes with special emphasis on bacterial control.Indian J Malariol. 1998 Dec;35(4):206-24. Indian J Malariol. 1998. PMID: 10748561 Review.
-
[Transgenic bioinsecticides inimical to parasites, but imical to environment].Wiad Parazytol. 2003;49(1):11-20. Wiad Parazytol. 2003. PMID: 16889013 Review. Polish.
References
-
- Berry C, O'Neil S, Ben-Dov E, Jones AF, Murphy L, Quail MA, Holden MT, Harris D, Zaritsky A, Parkhill J. 2002. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl Environ Microbiol 68:5082–5095. 10.1128/AEM.68.10.5082-5095.2002. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
