

Exploring the Immunogenicity of Noncanonical HLA-I Tumor Ligands Identified through Proteogenomics
- PMID: 36749875
- PMCID: PMC10261919
- DOI: 10.1158/1078-0432.CCR-22-3298
Exploring the Immunogenicity of Noncanonical HLA-I Tumor Ligands Identified through Proteogenomics
Abstract
Purpose: Tumor antigens are central to antitumor immunity. Recent evidence suggests that peptides from noncanonical (nonC) aberrantly translated proteins can be presented on HLA-I by tumor cells. Here, we investigated the immunogenicity of nonC tumor HLA-I ligands (nonC-TL) to better understand their contribution to cancer immunosurveillance and their therapeutic applicability.
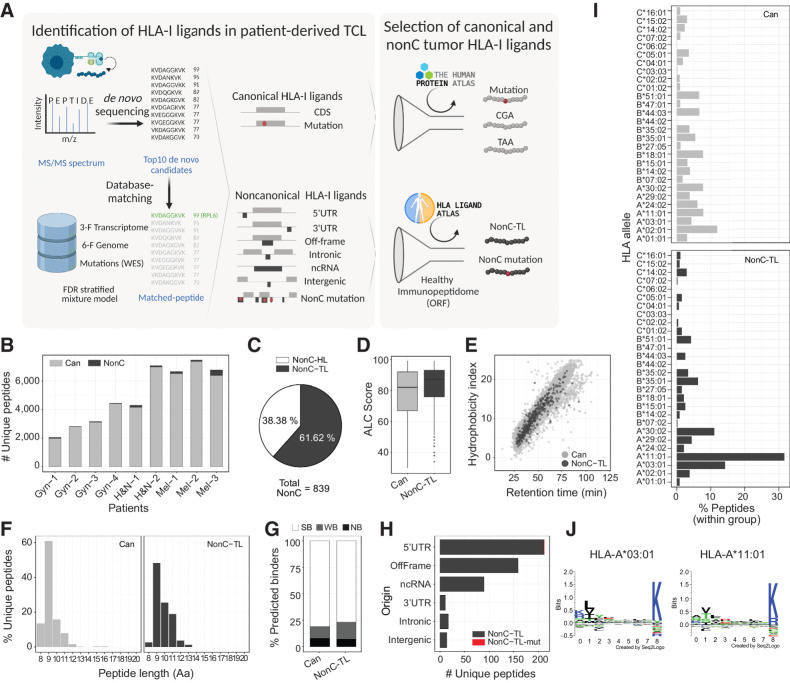
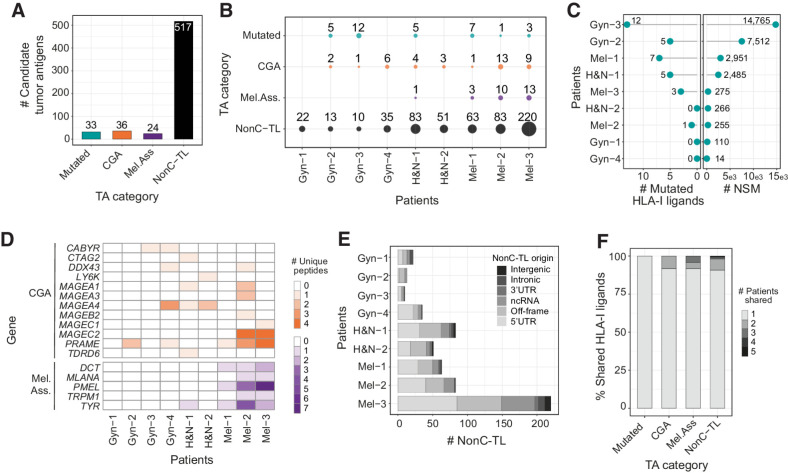
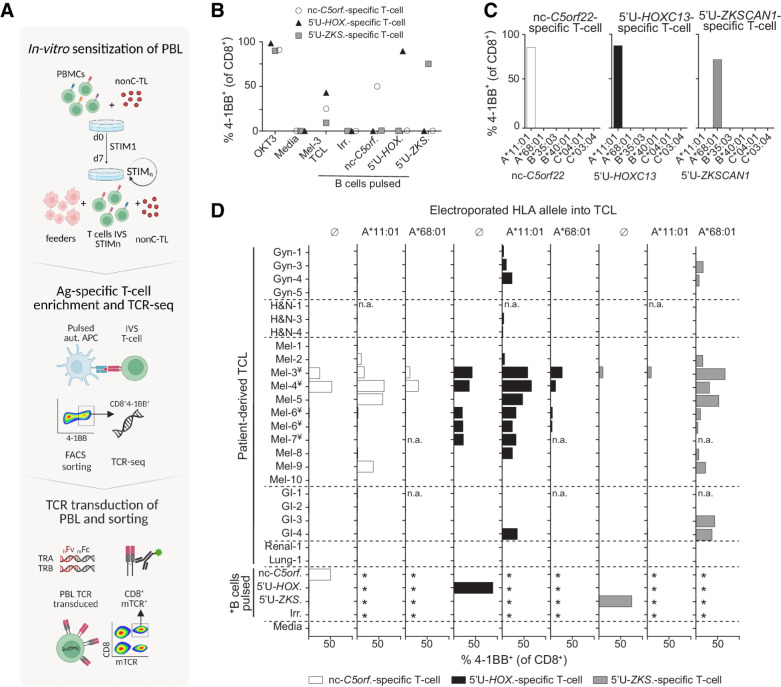
Experimental design: Peptides presented on HLA-I were identified in 9 patient-derived tumor cell lines from melanoma, gynecologic, and head and neck cancer through proteogenomics. A total of 507 candidate tumor antigens, including nonC-TL, neoantigens, cancer-germline, or melanocyte differentiation antigens, were tested for T-cell recognition of preexisting responses in patients with cancer. Donor peripheral blood lymphocytes (PBL) were in vitro sensitized against 170 selected nonC-TL to isolate antigen-specific T-cell receptors (TCR) and evaluate their therapeutic potential.
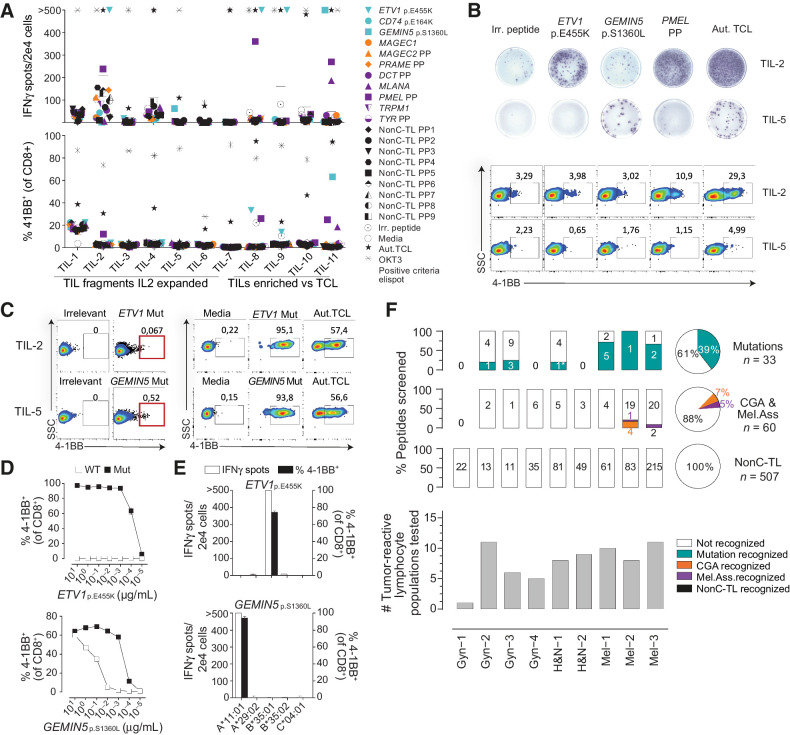
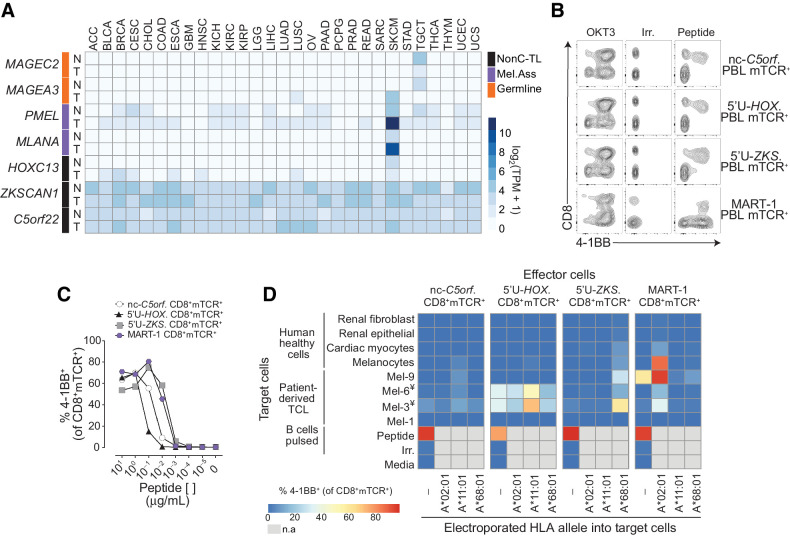
Results: We found no recognition of the 507 nonC-TL tested by autologous ex vivo expanded tumor-reactive T-cell cultures while the same cultures demonstrated reactivity to mutated, cancer-germline, or melanocyte differentiation antigens. However, in vitro sensitization of donor PBL against 170 selected nonC-TL, led to the identification of TCRs specific to three nonC-TL, two of which mapped to the 5' UTR regions of HOXC13 and ZKSCAN1, and one mapping to a noncoding spliced variant of C5orf22C. T cells targeting these nonC-TL recognized cancer cell lines naturally presenting their corresponding antigens. Expression of the three immunogenic nonC-TL was shared across tumor types and barely or not detected in normal cells.
Conclusions: Our findings predict a limited contribution of nonC-TL to cancer immunosurveillance but demonstrate they may be attractive novel targets for widely applicable immunotherapies. See related commentary by Fox et al., p. 2173.
©2023 The Authors; Published by the American Association for Cancer Research.
Figures
Comment in
-
Cancer's Dark Matter: Lighting the Abyss Unveils Universe of New Therapies.Clin Cancer Res. 2023 Jun 13;29(12):2173-2175. doi: 10.1158/1078-0432.CCR-23-0422. Clin Cancer Res. 2023. PMID: 37040070 Free PMC article.
Comment on
-
Cancer's Dark Matter: Lighting the Abyss Unveils Universe of New Therapies.Clin Cancer Res. 2023 Jun 13;29(12):2173-2175. doi: 10.1158/1078-0432.CCR-23-0422. Clin Cancer Res. 2023. PMID: 37040070 Free PMC article.
References
-
- Coulie PG, van den Eynde BJ, van der Bruggen P, Boon T. Tumor antigens recognized by T lymphocytes: at the core of cancer immunotherapy. Nat Rev Cancer 2014;14:135–46. - PubMed
-
- Brichard V, Pel Av, W6lfel T, W61fel C, de Plaen E, Leth6 B, et al. The tyrosinase gene codes for an antigen recognized by autologous cytolytic T lymphocytes on HLA-A2 melanomas. 1993. Available from: http://rupress.org/jem/article-pdf/178/2/489/1267835/489.pdf - PMC - PubMed
-
- van der Bruggen P, Traversari C, Chomez P, Lurquin C, de Plaen E, van den Eynde B, et al. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human. New Series 1991;254. - PubMed
