

Bacteriophages inhibit and evade cGAS-like immune function in bacteria
- PMID: 36750095
- PMCID: PMC9975087
- DOI: 10.1016/j.cell.2022.12.041
Bacteriophages inhibit and evade cGAS-like immune function in bacteria
Erratum in
-
Bacteriophages inhibit and evade cGAS-like immune function in bacteria.Cell. 2025 Sep 4;188(18):5118. doi: 10.1016/j.cell.2025.08.009. Epub 2025 Aug 22. Cell. 2025. PMID: 40848727 Free PMC article. No abstract available.
Abstract
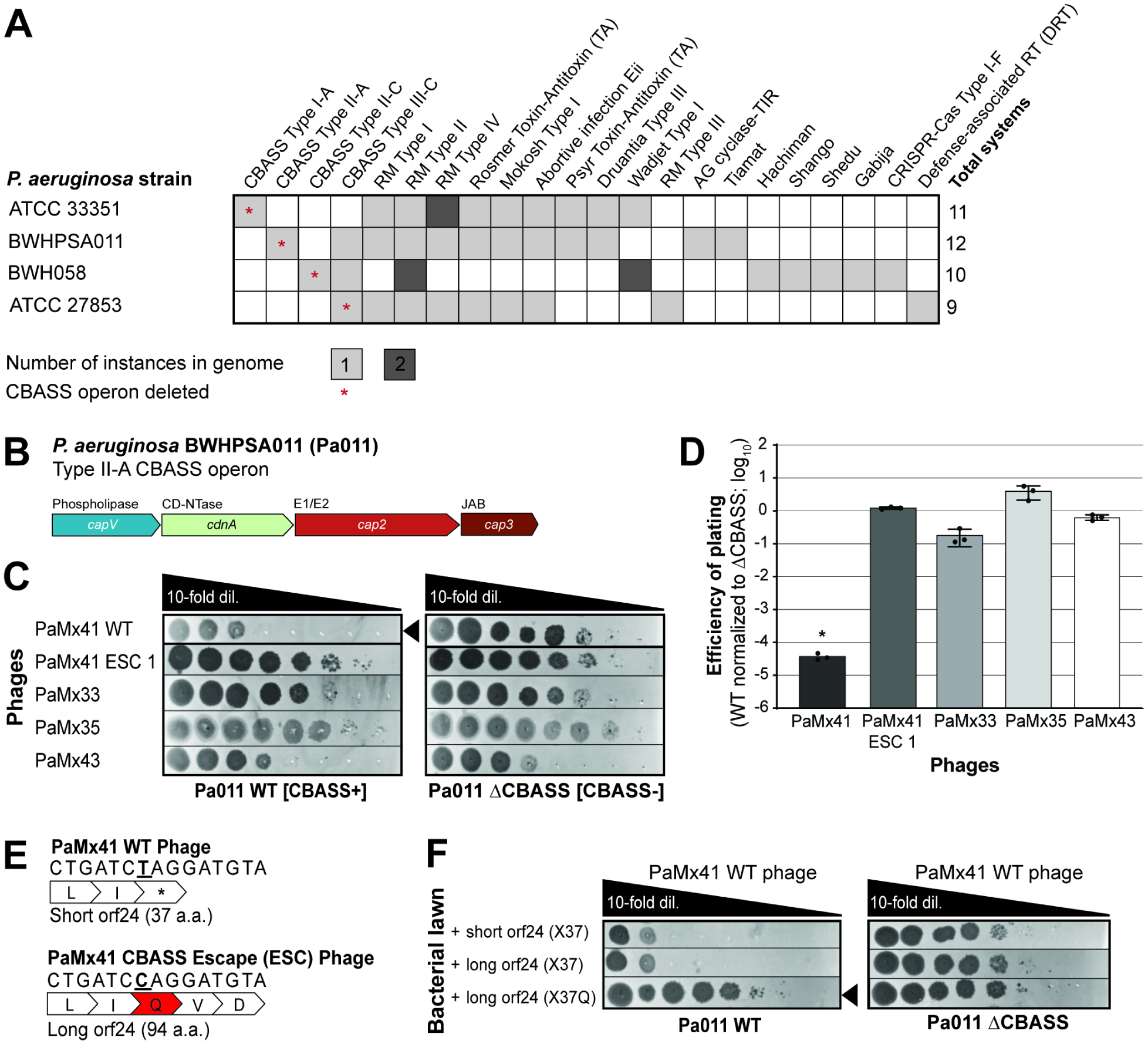
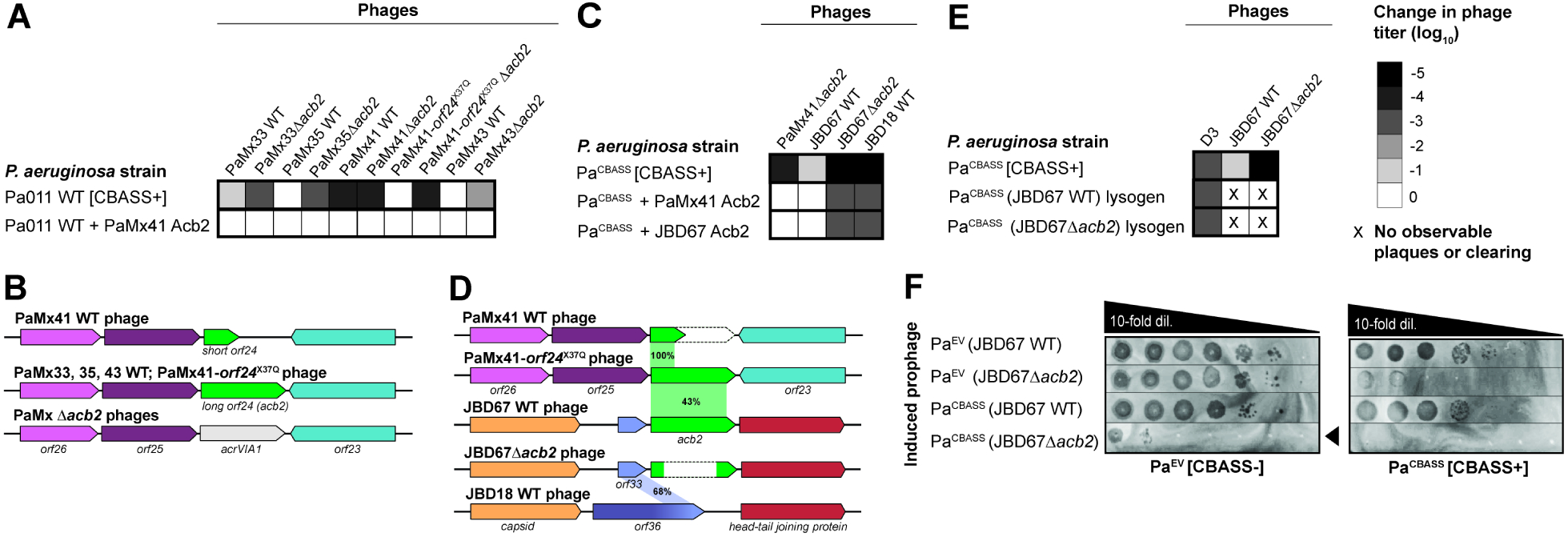
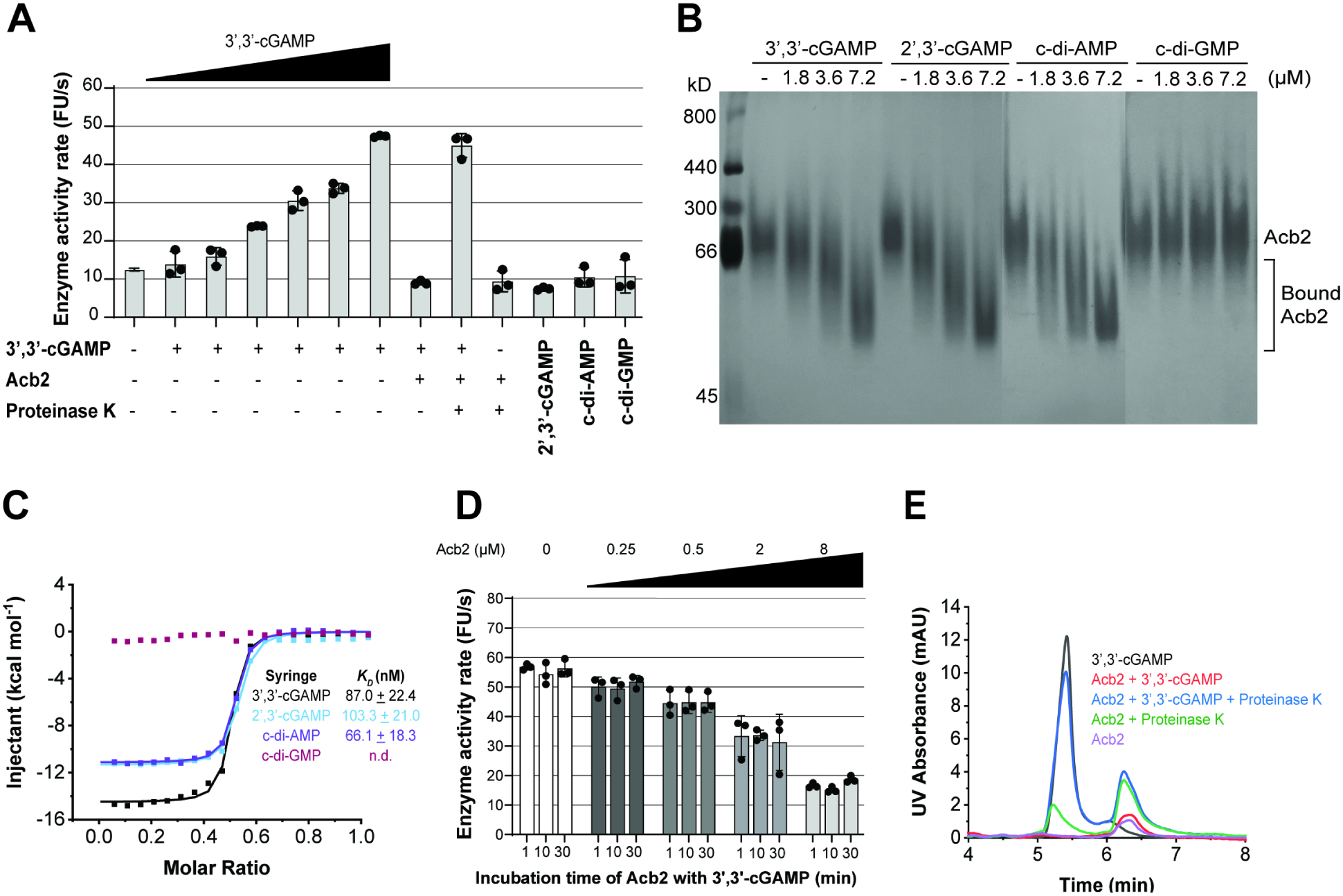
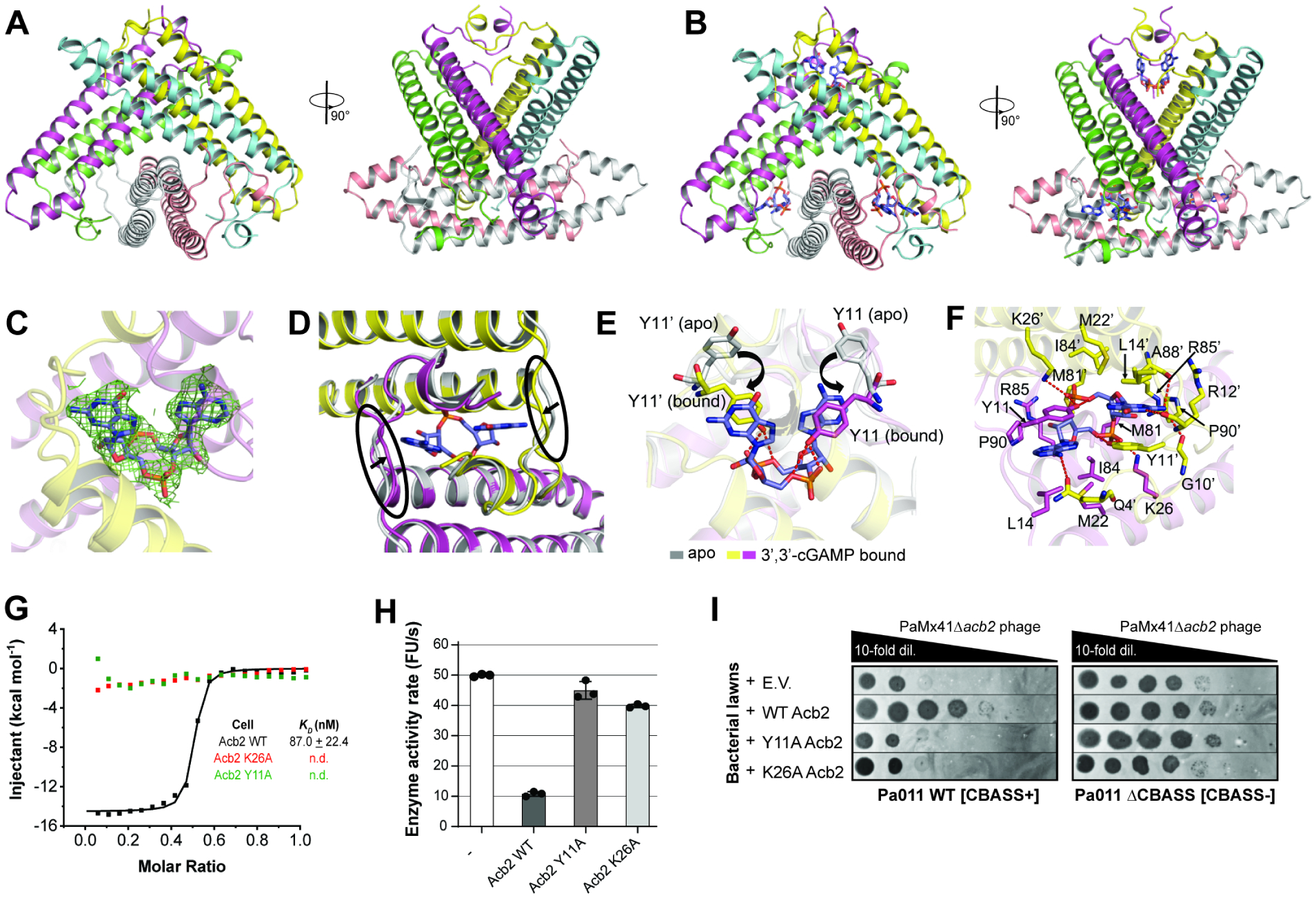
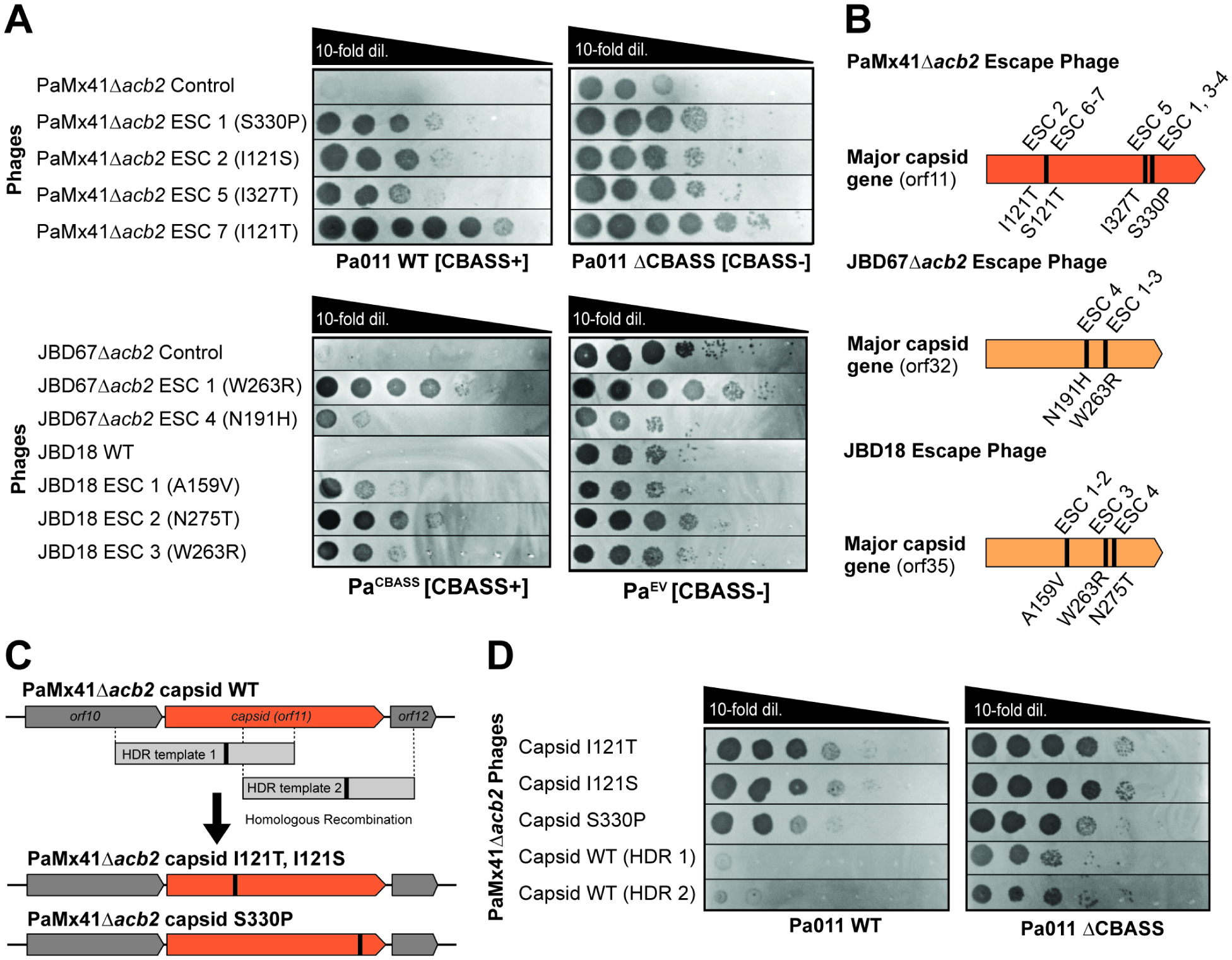
A fundamental strategy of eukaryotic antiviral immunity involves the cGAS enzyme, which synthesizes 2',3'-cGAMP and activates the effector STING. Diverse bacteria contain cGAS-like enzymes that produce cyclic oligonucleotides and induce anti-phage activity, known as CBASS. However, this activity has only been demonstrated through heterologous expression. Whether bacteria harboring CBASS antagonize and co-evolve with phages is unknown. Here, we identified an endogenous cGAS-like enzyme in Pseudomonas aeruginosa that generates 3',3'-cGAMP during phage infection, signals to a phospholipase effector, and limits phage replication. In response, phages express an anti-CBASS protein ("Acb2") that forms a hexamer with three 3',3'-cGAMP molecules and reduces phospholipase activity. Acb2 also binds to molecules produced by other bacterial cGAS-like enzymes (3',3'-cUU/UA/UG/AA) and mammalian cGAS (2',3'-cGAMP), suggesting broad inhibition of cGAS-based immunity. Upon Acb2 deletion, CBASS blocks lytic phage replication and lysogenic induction, but rare phages evade CBASS through major capsid gene mutations. Altogether, we demonstrate endogenous CBASS anti-phage function and strategies of CBASS inhibition and evasion.
Keywords: CBASS; Pseudomonas aeruginosa; acb; anti-CBASS; anti-phage immunity; bacteriophage; cGAS; capsid; escapers; innate immunity.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.B.-D. is a scientific advisory board member of SNIPR Biome, Excision Biotherapeutics, and LeapFrog Bio, and a scientific advisory board member and co-founder of Acrigen Biosciences. The Bondy-Denomy lab receives research support from Felix Biotechnology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
