

Structures of LRP2 reveal a molecular machine for endocytosis
- PMID: 36750096
- PMCID: PMC9993842
- DOI: 10.1016/j.cell.2023.01.016
Structures of LRP2 reveal a molecular machine for endocytosis
Abstract
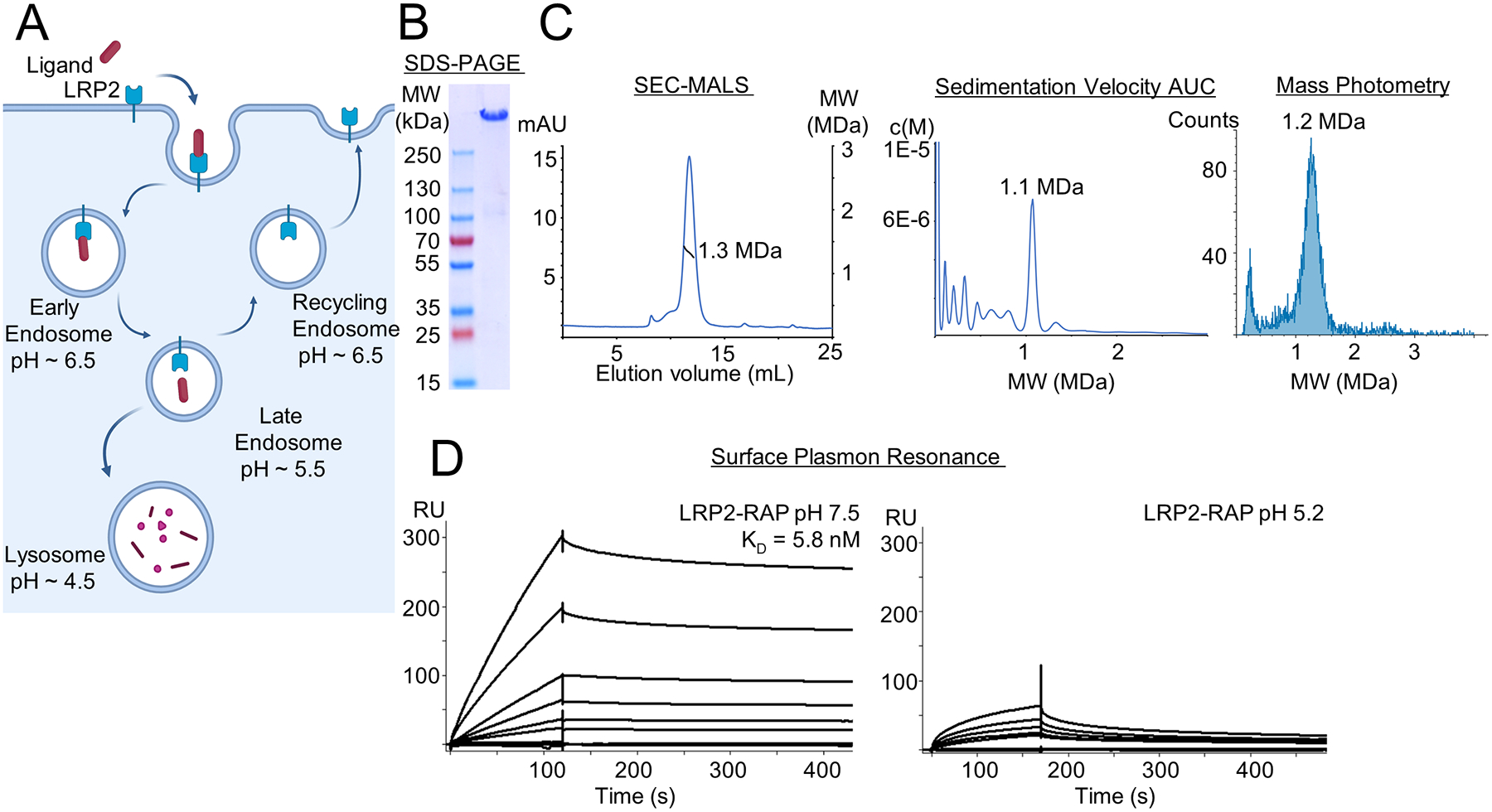
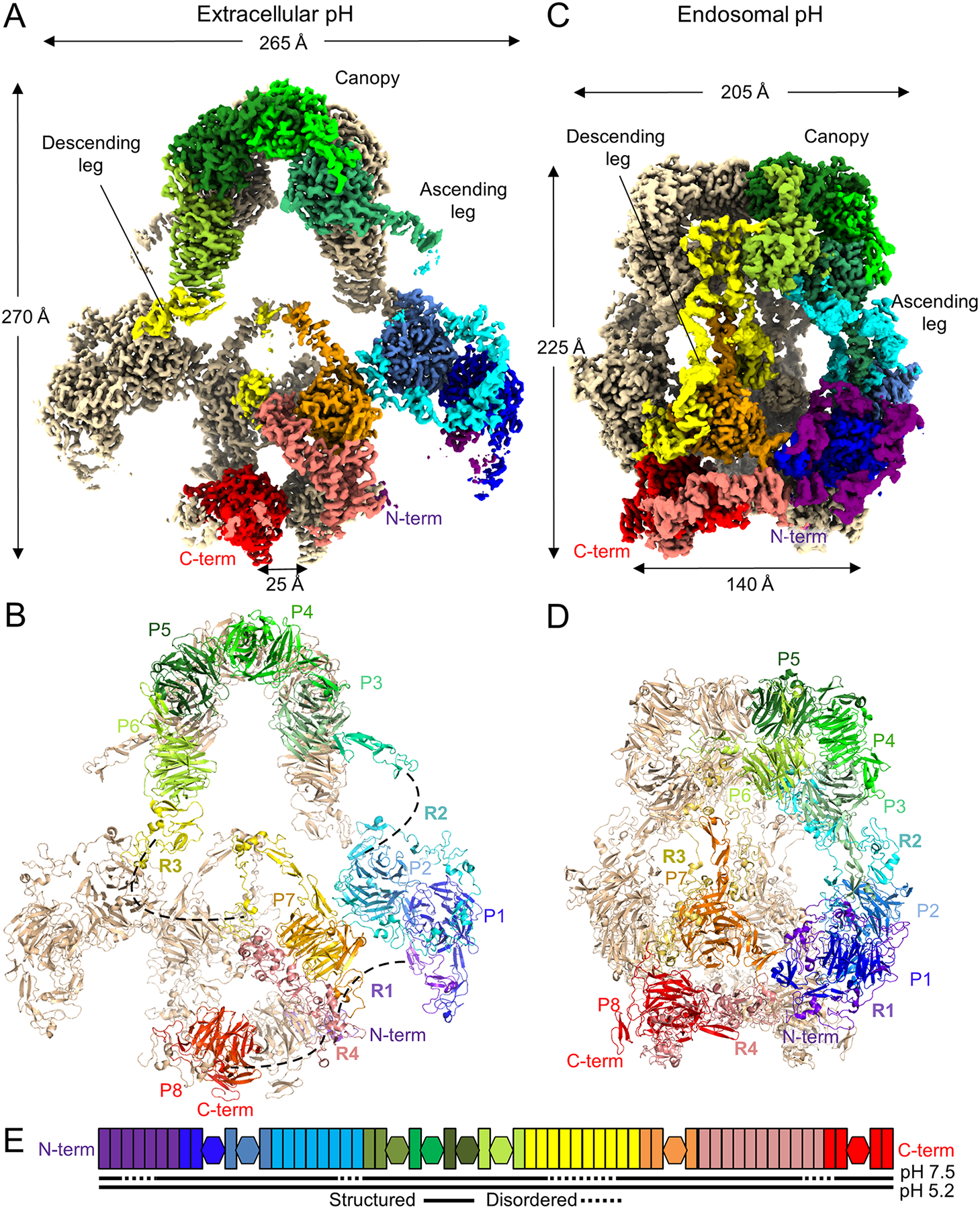
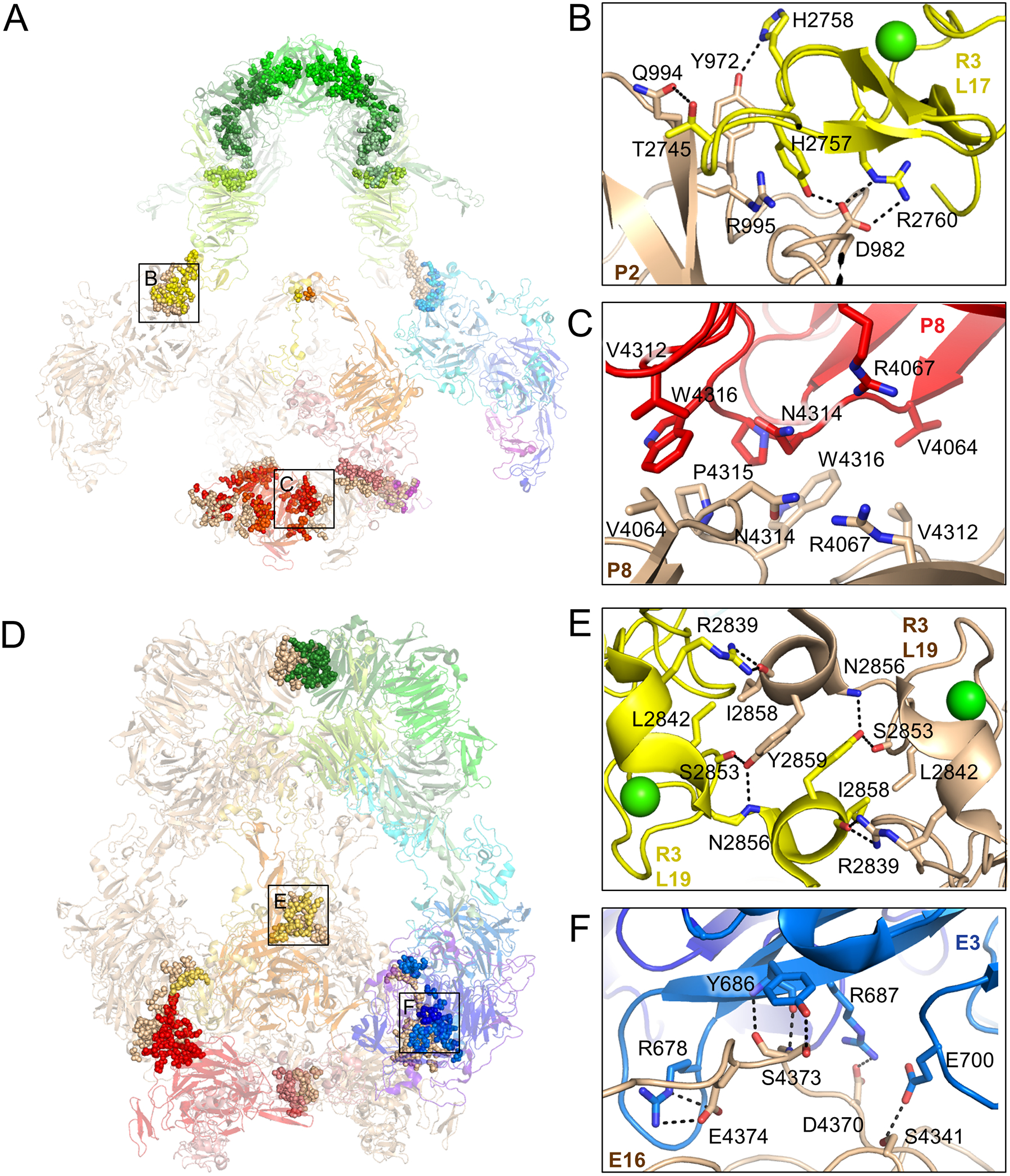
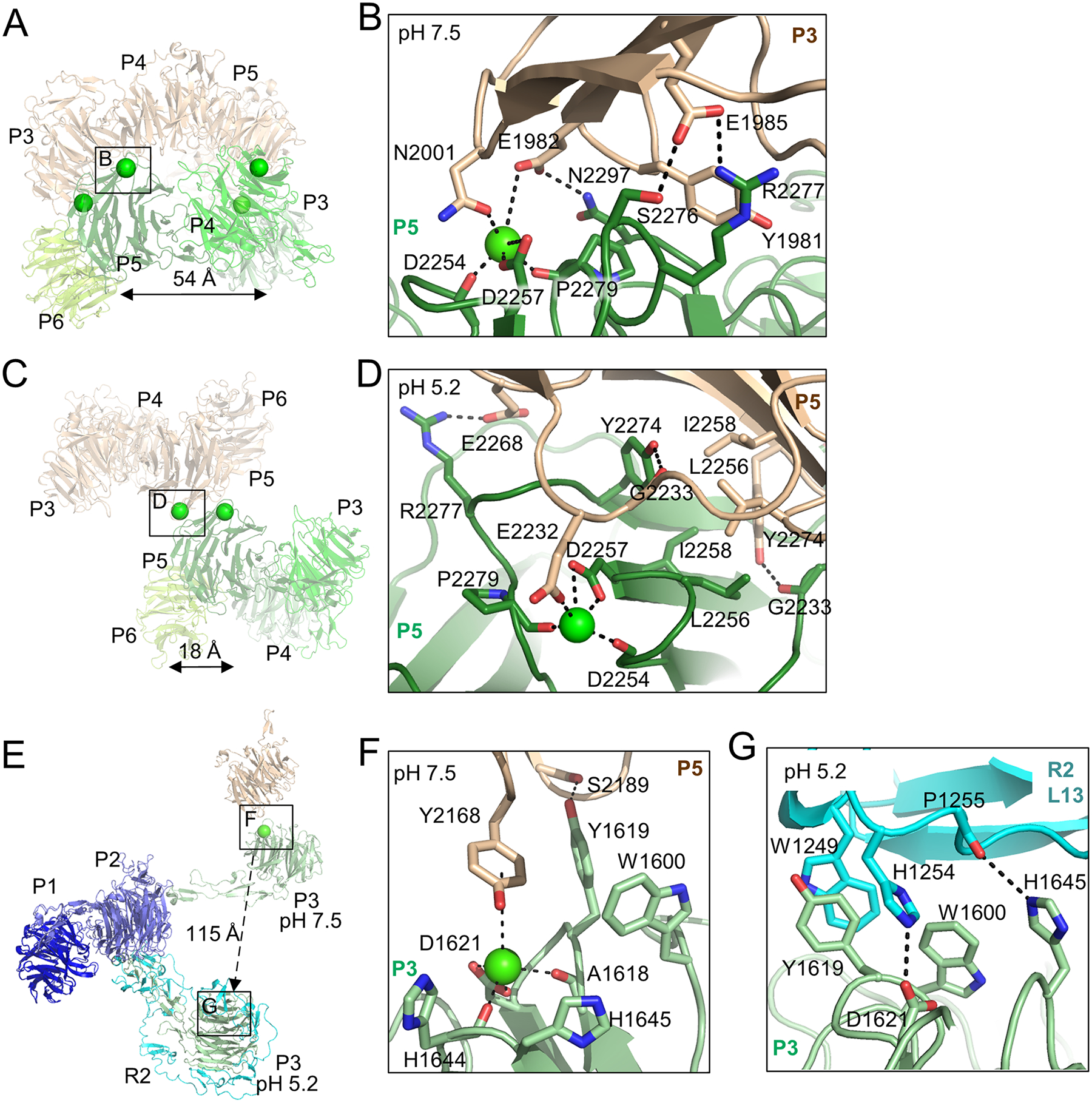
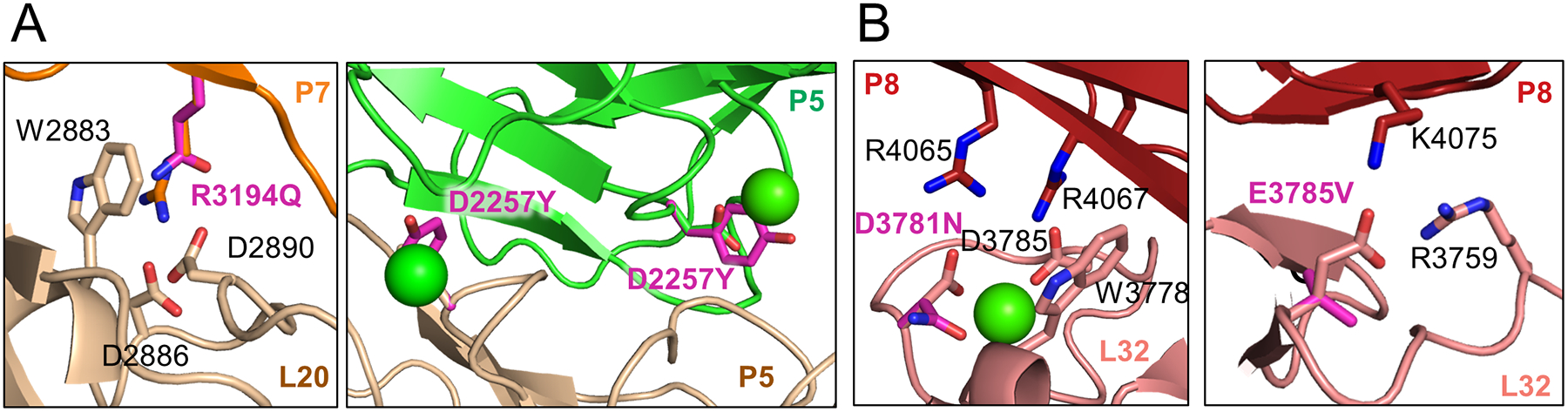
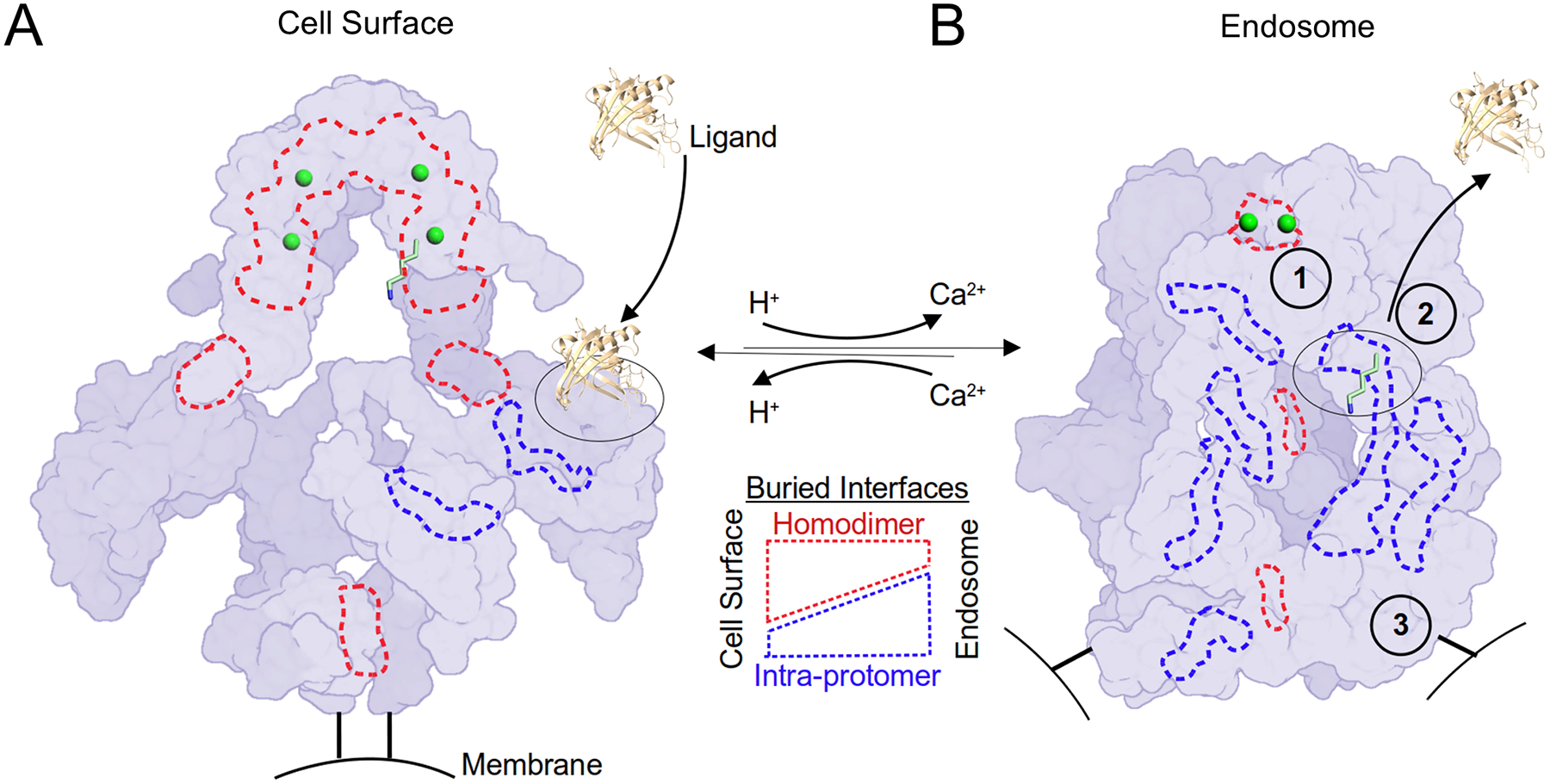
The low-density lipoprotein (LDL) receptor-related protein 2 (LRP2 or megalin) is representative of the phylogenetically conserved subfamily of giant LDL receptor-related proteins, which function in endocytosis and are implicated in diseases of the kidney and brain. Here, we report high-resolution cryoelectron microscopy structures of LRP2 isolated from mouse kidney, at extracellular and endosomal pH. The structures reveal LRP2 to be a molecular machine that adopts a conformation for ligand binding at the cell surface and for ligand shedding in the endosome. LRP2 forms a homodimer, the conformational transformation of which is governed by pH-sensitive sites at both homodimer and intra-protomer interfaces. A subset of LRP2 deleterious missense variants in humans appears to impair homodimer assembly. These observations lay the foundation for further understanding the function and mechanism of LDL receptors and implicate homodimerization as a conserved feature of the LRP receptor subfamily.
Keywords: CD91; LDL; LRP1; LRP2; cryo-EM; endocytosis; megalin; pH-sensitive; proteinuria; recycling.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Comment in
-
The structure of megalin: shedding new light on receptor-mediated endocytosis.Kidney Int. 2024 Jan;105(1):11-14. doi: 10.1016/j.kint.2023.05.026. Epub 2023 Jun 26. Kidney Int. 2024. PMID: 37380129 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
